能夠結合組織因子途徑抑制劑(1‑161)上的兩個表位的抗體的製作方法
2023-11-11 20:52:03 2
名為「140072 SEQUENCE LISTING_ST25」的序列表(48KB)創建於2015年9月16日,並通過引用併入本文。
發明領域
本發明涉及能夠同時結合組織因子途徑抑制劑(TFPI)的N-末端區域(殘基1-161)內的兩個表位的雙特異性抗體及其組合物。本發明還涉及這類抗體的製藥和治療用途。
背景
在出血的個體中,當血管外TF暴露於血液中的FVIIa時,組織因子/因子VIIa(TF/FVIIa)複合物引發凝血。TF/FVIIa複合物的形成導致因子X(FX)活化為FXa,該FXa與活化的因子V(FVa)一起生成有限量的凝血酶。小量凝血酶會激活血小板,這繼而導致血小板磷脂的表面暴露,該血小板磷脂支持由活化的因子VIII(FVIIIa)和因子IX(IXa)組成的tenase複合物的裝配和結合。tenase複合物是非常有效的FX活化催化劑,並且在該第二步中生成的FXa充當引起最終凝血酶爆發的FVa/FXa凝血酶原酶(pro-thrombinase)複合物中的活性蛋白酶。凝血酶切割纖維蛋白原產生纖維蛋白單體,該纖維蛋白單體聚合而形成密封滲漏的血管並阻止出血的纖維蛋白網絡。快速且廣泛的凝血酶爆發是形成固態和穩定的纖維蛋白凝塊的先決條件。
由FVIII或FIX缺乏引起的FXa和凝血酶生成上不充分的放大分別是A型血友病和B型血友病患者中出血素質的潛在原因。在血友病患者中,FXa生成主要由TF/FVIIa複合物驅動,因為FVIII或FIX缺乏僅導致通過tenase複合物的不完全的FXa生成。然而,TF/FVIIa介導的FX向FXa的活化是暫時的,因為組織因子途徑抑制劑(TFPI)在自調節環路中抑制因子Xa和TF/FVIIa複合物。反饋抑制導致TF/FVIIa/FXa/TFPI複合物的形成。在凝血啟動過程中,中和TFPI抑制使TF/FVIIa介導的FX活化延長,從而促進由於tenase活性受損而導致FXa生成不足的血友病患者中的止血,該tenase活性受損是由例如FVIII或FIX缺乏引起的。
在凝血啟動後,TF/FVIIa介導的FXa生成被TFPI緊密下調。TFPI是一種緩慢的緊密結合的競爭性抑制劑,其通過抑制TF-FVIIa和FXa兩者來調節FX活化和活性。當FXa與TF/FVIIa複合物結合或在其附近結合到膜上時,TF/FVIIa在作為限速步驟涉及FXa的TFPI抑制的過程中被TFPI抑制(Baugh等人,1998,JBC,273:4378-4386)。FXa的TFPI抑制發生在兩相反應中,該反應最初導致鬆散的TFPI-FXa複合物,該複合物緩慢地重排成緊密結合的TFPI-FXa複合物,其中TFPI的第二Kunitz型抑制劑結構域(KPI-2)結合併阻斷FXa的活性位點。TFPI的第一Kunitz型抑制劑結構域(KPI-1)有助於緊密TFPI-FXa複合物的形成,並且其直接結合併阻斷TF結合的FVIIa的活性位點。
能夠結合TFPI的抗體是本領域已知的。例如,WO2010/072691、WO2012/001087和WO2012/135671公開了單克隆抗體(mAb),其中每一種均能夠與TFPI的一個特異性表位結合。以下限制可能適用於靶向單個TFPI表位(例如KPI結構域上的)的這類抗體,這通常局限於由單個可變區限定的抗體的互補位區域。首先,TF/FVIIa/FXa複合物的最終抑制依賴於分散在TFPI上的互補區域與TF/FVIIa/FXa複合物之間的幾種相互作用。這不僅適用於TFPI的KPI-1和KPI-2分別與FVIIa和FXa的活性位點的直接結合,還適用於與TF/FVIIa/FXa外結合位點(exosite)的相互作用,該相互作用涉及TFPI的N-和C-末端區的區域。例如結合單個KPI的單克隆抗體可能不能完全阻斷TFPI的所有抑制功能,特別是在生理上升高的TFPI濃度下。其次,單特異性抗體的效力可能受到抗體親和力和/或TPFI分子柔性的阻礙,這兩者都將需要高劑量的前者。第三,由於TFPI-mAb複合物的腎清除率降低,或由於因TFPI-mAb複合物形成而減弱的其它清除機制,用單克隆抗體靶向TFPI可能會導致TFPI在循環中積累。
本發明人預期本文公開的雙特異性抗體可以解決這些限制。
技術實現要素:
本發明涉及一種能夠特異性結合人TFPI(SEQ ID NO:1)的位置1至161內的第一表位和第二表位的雙特異性抗體。
在一個方面,該雙特異性抗體的第一表位可在人TFPI的位置1-96內,如位置1-76內,如Kunitz型抑制劑1(KPI-1)結構域(殘基26-76)內。該KPI-1表位可包含SEQ ID NO:1的胺基酸殘基Arg 41、Arg 65和/或Glu 67中的一個或多個,並且可進一步包含SEQ ID NO:1的胺基酸殘基Leu 16、Pro 17、Leu 19、Lys 20、Leu 21、Met 22、Phe 25、Cys 35、Ala 37、Met 39、Tyr 56、Gly 57、Gly 58、Cys 59、Glu 60、Gly 61、Asn 62、Gln 63、Phe 66、Glu 71和Met 75中的一個或多個。
在一個方面,該雙特異性抗體的第二表位可在人TFPI的位置77-161內,如位置97-161內,如Kunitz型抑制劑2(KPI-2)結構域(殘基97-147)內。該KPI-2表位可包含SEQ ID NO:1的胺基酸殘基Arg 107,並且可進一步包含SEQ ID NO:1的胺基酸殘基Glu 100、Glu 101、Asp 102、Pro 103、Tyr 109、Thr 111、Tyr 113、Phe 114、Asn 116、Gln 118、Gln 121、Cys 122、Glu 123、Arg 124、Phe 125、Lys 126和Leu 140中的一個或多個。
本發明的雙特異性抗體可以是IgG形式,如全長IgG形式,或者可以是兩個抗體片段的化學綴合物,如兩個Fab片段或scFv片段的綴合物,或其組合。該雙特異性抗體優選是人抗體或人源化抗體。
本發明還涉及一種包含根據本發明的雙特異性抗體和藥學上可接受的載體的藥物組合物。所述雙特異性抗體或包含該抗體的組合物可用作藥物。所述藥物可用於治療先天性、獲得性和/或醫源性凝血障礙,如具有或不具有抑制物的A型血友病,以及具有或不具有抑制物的B型血友病。
附圖簡述
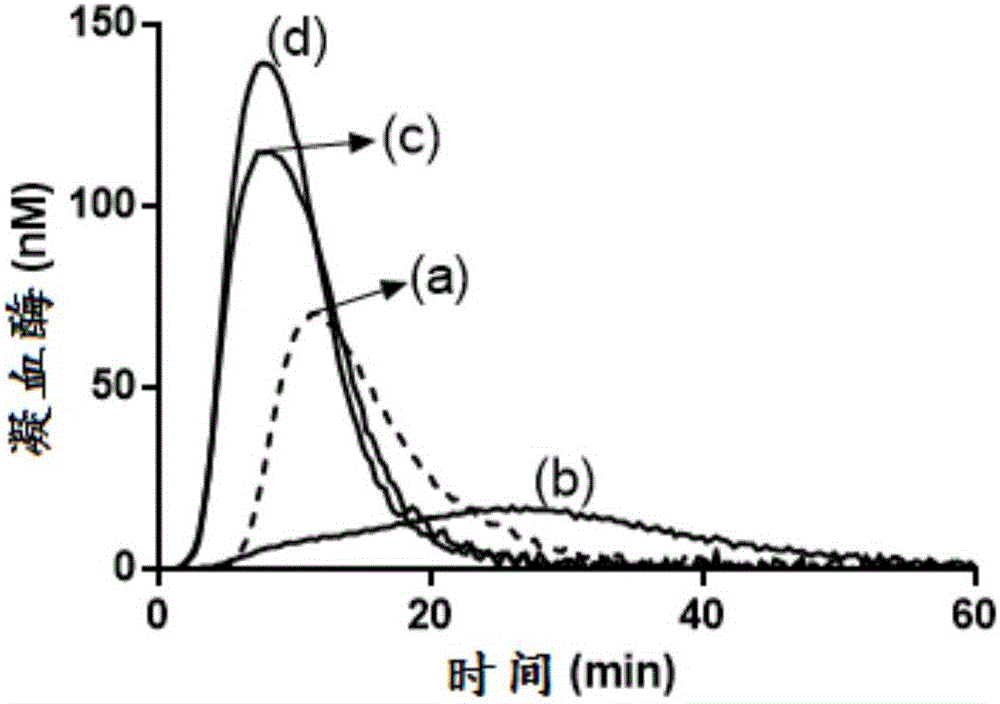
圖1:在TFPI水平升高的A型血友病樣條件下的人血漿中,某些TFPI(1-79)抗體及TFPI KPI-2抗體mAb 2021增強了凝血酶生成。曲線(a)顯示了在沒有另外添加的情況下於正常人血漿中獲得的結果。曲線(b)顯示了通過添加100μg/ml FVIII抗體(HTI PAHFVIII-S)和20nM全長TFPIα獲得A型血友病樣條件時的結果。由20nM TFPIα聯合FVIII中和導致的凝血酶生成的抑制不受添加200nM TFPI(1-79)抗體mAB 1F91(c)的影響。200nM TFPI(1-79)抗體mAb 2F22(d)和KPI-2抗體mAb 2021(e)的添加確實逆轉了凝血酶的生成,但沒有使其完全恢復至在沒有FVIII抗體的情況下於正常人中獲得的水平(曲線(a))。
圖2:在TFPI水平升高的A型血友病樣條件下的人血漿中,通過將TFPI(1-79)與TFPI-KPI-2抗體組合大大增強了凝血酶生成。曲線(a)顯示了在沒有另外添加的情況下於正常人血漿中獲得的結果。曲線(b)顯示了通過添加100μg/ml FVIII抗體(HTI PAHFVIII-S)、20nM TFPIα和200nM KPI-2抗體mAb 2021獲得A型血友病樣條件時的結果。通過將100nM mAb 2021與100nM TFPI(1-79)抗體mAb 1F91(c)或mAb 2F22(d)組合,(b)中採用200nM mAb 2021的不完全中和得到逆轉。(c)和(d)中TFPI(1-79)與KPI-2抗體的組合導致了比正常人血漿(a)更高的凝血酶峰值。
圖3:來自TFPI FL與被捕獲的雙特異性抗體mAb 0421和BiFab 9041結合的結果。
序列簡述
SEQ ID NO:1代表人TFPIα的胺基酸序列。
SEQ ID NO:2代表TFPI(1-161)的胺基酸序列。
SEQ ID NO:3代表標記的跨越KPI-2的TFPI片段的胺基酸序列。
SEQ ID NO:4代表單克隆抗體(mAb)2F3的可變重鏈的胺基酸序列。
SEQ ID NO:5代表單克隆抗體(mAb)2F3的可變輕鏈的胺基酸序列。
SEQ ID NO:6代表單克隆抗體(mAb)2F22的可變重鏈的胺基酸序列。
SEQ ID NO:7代表單克隆抗體(mAb)2F22的可變輕鏈的胺基酸序列。
SEQ ID NO:8代表單克隆抗體(mAb)2F45的可變重鏈的胺基酸序列。
SEQ ID NO:9代表單克隆抗體(mAb)2F45的可變輕鏈的胺基酸序列。
SEQ ID NO:10代表單克隆抗體(mAb)1F91的可變重鏈的胺基酸序列。
SEQ ID NO:11代表單克隆抗體(mAb)1F91的可變輕鏈的胺基酸序列。
SEQ ID NO:12代表用於克隆抗體重鏈的引物的核酸序列。
SEQ ID NO:13代表用於克隆抗體輕鏈的引物的核酸序列。
SEQ ID NO:14代表Fab 0296的截短的鼠-人嵌合重鏈的胺基酸序列。
SEQ ID NO:15代表單克隆抗體mAb 0294、Fab 0296、Fab 0295和mAb 0336的鼠-人嵌合輕鏈的胺基酸序列。
SEQ ID NO:16代表標記的TFPI KPI-1/N-末端(TFPI(1-79))的胺基酸序列。
SEQ ID NO:17代表單克隆抗體mAb 0294的鼠-人嵌合重鏈的胺基酸序列。
SEQ ID NO:18代表Fab 0295的截短的鼠-人嵌合重鏈的胺基酸序列。
SEQ ID NO:19代表mAb 2021的重鏈可變域(VH)的胺基酸序列。
SEQ ID NO:20代表mAb 2021的輕鏈可變域(VL)的胺基酸序列。
SEQ ID NO:21代表Fab 0094的重鏈(HC)的胺基酸序列。
SEQ ID NO:22代表Fab 0094的輕鏈(LC)的胺基酸序列。
SEQ ID NO:23代表Fab 0088的重鏈(HC)的胺基酸序列。
SEQ ID NO:24代表Fab 0088的輕鏈(LC)的胺基酸序列。
SEQ ID NO:25代表Fab 0313的重鏈(HC)的胺基酸序列。
SEQ ID NO:26代表Fab 0313的輕鏈(LC)的胺基酸序列。
SEQ ID NO:27代表mAb 0310的重鏈(HC)的胺基酸序列。
SEQ ID NO:28代表mAb 0310的輕鏈(LC)的胺基酸序列。
SEQ ID NO:29代表mAb 0336的重鏈(HC)的胺基酸序列。
詳述
在體內,組織因子途徑抑制劑(TFPI)存在於若干區室中。TFPI中的大部分與血管內皮相關聯,少部分在血液中循環。已經描述了人類中的兩種TFPI剪接變體:TFPIα和TFPIβ。TFPIβ可能是在內皮細胞表面上表達的主要形式,而TFPIα的胞內儲存可經某些刺激被釋放至循環中。TFPIα作為全長或截短的蛋白質在血液中循環,與脂蛋白或血小板相關聯。
成熟人TFPIα是由酸性N-末端區域、由連接區穿插的三個串聯排列的Kunitz型抑制劑結構域(KPI-1、KPI-2和KPI-3)和鹼性C-末端尾部組成的276個胺基酸的蛋白質(SEQ ID NO:1)。KPI-1、KPI-2和KPI-3結構域分別被定義為SEQ ID NO:1的殘基26-76、殘基97-147和殘基189-239。成熟人TFPIβ是經由糖基磷脂醯肌醇(GPI)-錨共價連接至內皮細胞表面的193個胺基酸的蛋白質。TFPIβ的前161個胺基酸與TFPIα相同(對應於SEQ ID NO:1的殘基1-161),而C-末端序列的最後12個胺基酸與TFPIα不相關,並且具有連接至殘基193的GPI-錨。
本發明涉及結合TFPI的殘基1-161內的兩個不同和/或獨特表位的雙特異性抗體。各自具有獨特抗原識別位點的兩個單特異性抗體形成本發明雙特異性抗體的抗原結合片段或「臂」的基礎。該雙特異性抗體的第一抗原結合片段(或「臂」)針對SEQ ID NO:1的胺基酸殘基1-96內的表位;即,針對位於包含N末端、KPI-1結構域(SEQ ID NO:1的殘基26-76)以及KPI-1與KPI-2結構域之間的連接區的區域內的表位。所述表位優選位於包含TFPI的胺基酸1-76的區域內。該雙特異性抗體的第二抗原結合片段(或「臂」)針對SEQ ID NO:1的胺基酸77-161內的表位;即,包含KPI-1與KPI-2結構域之間的連接區、KPI-2結構域以及KPI-2與KPI-3結構域之間的連接區的TFPI區域。所述表位優選位於TFPI的KPI-2結構域(SEQ ID NO:1的殘基97-147)內。
能夠在SEQ ID NO:1的胺基酸77-161內,如在KPI-2結構域內結合的雙特異性抗體的單臂本身可如同單特異性抗體一樣,不能顯著阻止TFPI的抑制活性。然而,當該雙特異性抗體的另一臂與包含TFPI的胺基酸1-76的區域如TFPI的KPI-1結構域結合時,所述單臂可更顯著地促進TFPI抑制的特異性阻斷,反之亦然。
優選地,如本文所公開的雙特異性KPI-1/KPI-2結合抗體的主要結合模式是分子內的,即其中該雙特異性KPI-1/KPI-2結合抗體的兩個臂與單個TFPI分子結合;而不是分子間的,即其中該雙特異性KPI-1/KPI-2結合抗體的兩個臂與不同的TFPI分子結合。分子內結合形成1∶1TFPI抗體複合物,而分子間結合模式可導致不希望的TFPI和雙特異性抗體的較大裝配體的形成。可通過降低其中一個抗體臂如KPI-1結合臂的親和力來刺激分子內結合模式。
當比較本發明雙特異性抗體的促凝血作用與衍生出該抗體的單特異性抗體的預期累加作用時,前者可具有優異的表現。例如,即使當TFPI的濃度相對於例如正常生理水平升高時,該雙特異性抗體也可具有阻斷TFPI所有抑制功能的能力。為了獲得雙特異性抗體如Fab-Fab綴合物(本文也稱為BiFab),針對本發明兩個不同表位的兩個抗原結合片段的連接,由於親合力效應,可導致比通過這兩個不同抗原結合部分的組合效應所獲得的更強的TFPI活性中和。
本發明的雙特異性抗體可具有調節所有TFPI集合的活性的能力。
如本文中所使用的術語「TFPI」包括可來源於任何合適生物體的天然存在形式的組織因子途徑抑制劑(TFPI)。例如,如本文所述使用的TFPI可以是哺乳動物TFPI,如人、小鼠、大鼠、靈長類、牛、羊、兔或豬TFPI。優選地,該TFPI是人TFPI。該TFPI可以是成熟形式的TFPI,如在合適的細胞內經歷翻譯後加工的TFPI蛋白質。這樣的成熟TFPI蛋白質例如可被糖基化。該TFPI可以是全長TFPI蛋白質。術語TFPI還包括這樣的TFPI分子的變體、同種型和其它同源物。TFPI活性是指其抑制活性。變異TFPI分子的特徵通常在於具有與天然存在的TFPI相同類型的活性,如中和FXa的催化活性的能力或抑制TF-FVIIa/FXa複合物的能力。
本文中的術語「抗體」是指自免疫球蛋白序列衍生的蛋白質,其能夠與抗原或其一部分特異性結合。術語抗體包括但不限於任何類別(或同種型)的全長抗體,即IgA、IgD、IgE、IgG、IgM和/或IgY。該術語也可包括全長抗體的一個或多個抗原結合片段。與抗原或其一部分特異性結合的抗體可僅與該抗原或其一部分結合,或者其可以與有限數目的同源性抗原或其部分結合。
天然的全長抗體通常包含至少四條多肽鏈:通過二硫鍵連接的兩條重(H)鏈和兩條輕(L)鏈。在一些情況下,天然抗體包含少於四條鏈,如在駱駝科動物中發現的僅含重鏈的抗體(VHH片段)和在軟骨魚類(Chondrichthyes)中發現的IgNAR的情況。藥學上特別感興趣的一類免疫球蛋白是IgG。在人類中,根據其重鏈恆定區的序列,IgG類別可被細分為四個亞類:IgG1、IgG2、IgG3和IgG4。根據其序列組成的差異,輕鏈可被分為兩種類型:κ和λ鏈。IgG分子由兩條通過兩個或更多個二硫鍵相互連接的重鏈和兩條各自通過二硫鍵連接至重鏈的輕鏈組成。IgG重鏈可包含一個重鏈可變區(VH)和最多三個重鏈恆定(CH)區:CH1、CH2和CH3。輕鏈可包含輕鏈可變區(VL)和輕鏈恆定區(CL)。VH和VL區可進一步細分為被稱為互補決定區(CDR)或高變區(HvR)的高變性區域,其中散布有被稱為框架區(FR)的更加保守的區域。VH和VL區通常由三個CDR和四個FR組成,它們從氨基末端到羧基末端以如下順序排列:FR1、CDR1、FR2、CDR2、FR3、CDR3、FR4。重鏈和輕鏈的具有高變區的可變域構成能夠與抗原相互作用的結構域,而抗體的恆定區可以介導免疫球蛋白與宿主組織或因子的結合,該宿主組織或因子包括但不限於免疫系統的各種細胞(效應細胞)、Fc受體和經典補體系統的C1複合物的第一組分(C1q)。
如本文所使用的,術語「單特異性抗體」是指具有單一抗原識別位點(即,為單價)或各自對一個共同靶抗原具有特異性的兩個相同抗原識別位點(即,為二價)的抗體。
本發明的單特異性抗體可以是單克隆抗體,從這種意義上講,它們代表由單個B細胞或由B細胞的克隆群表達的一組獨特的重鏈和輕鏈可變域序列。本發明的抗體可採用本領域技術人員已知的各種方法來生產和純化。例如,抗體可由雜交瘤細胞產生。抗體可通過B細胞擴增來產生。抗體或其片段可以在哺乳動物或微生物表達系統中或通過體外翻譯重組表達。抗體或其片段還可通過例如噬菌體展示、細菌展示、酵母展示、哺乳動物細胞展示或核糖體或mRNA展示重組表達為細胞表面結合分子。一旦產生,可以針對其結合TFPI(1-161)、全長TFPIα和TFPIβ,如人TFPI(1-161)、全長人TFPIα和人TFPIβ的能力篩選抗體。
抗體的抗原結合片段也可以是根據本發明的單特異性抗體,因為已經證明,抗體的抗原結合功能可由全長抗體的片段來行使。術語抗體的「抗原結合片段」是指一個或多個保留特異性結合或識別抗原的能力的抗體片段,該抗原例如是TFPIα如人TFPIα(SEQ ID NO:1),如人TFPI(1-161)(SEQ ID NO:2),如人TFPI KPI-2構建體(SEQ ID NO:3),或另一種靶分子,如本文所述。抗原結合片段的例子包括Fab、Fab′、Fab2、Fab′2、FabS、Fv(一般為抗體單臂的VL和VH域)、單鏈Fv(scFv;參見例如Bird等人,Science(1988)242:42S-426;和Huston等人PNAS (1988)85:5879-5883)、dsFv、Fd(一般為VH和CH1域)和dAb(一般為VH域)片段;VH、VL、VhH和V-NAR域;包含單條VH和單條VL鏈的單價分子;微體(minibodies)、雙抗體(diabodies)、三抗體(triabodies)、四抗體(tetrabodies)和κ體(kappa bodies)(參見例如Ill等人Protein Eng(1997)10:949-57);駱駝IgG;IgNAR;以及一個或多個分離的CDR或功能互補位,其中分離的CDR或抗原結合殘基或多肽可以締合或連接在一起,從而形成功能抗體片段。各種類型的抗體片段已在例如Holliger和Hudson,Nat Biotechnol(2005)23:1126-1136、WO2005040219以及公開的美國專利申請20050238646和20020161201中描述或綜述。這些抗體片段可採用本領域技術人員已知的常規技術獲得,並且可以以與完整抗體相同的方式針對應用性篩選這些片段。
抗體的「Fab片段」,包括「Fab」、「Fab」』和「Fab』2」片段,可通過在連接抗體重鏈的鉸鏈半胱氨酸殘基的N-末端或C-末端側上的鉸鏈區中切割重鏈而由所述抗體得到。「Fab」片段包括輕鏈的可變域和恆定域以及重鏈的可變域和第一個恆定域(CH1)。「Fab′2」片段包含一對通常通過其鉸鏈半胱氨酸共價連接的「Fab」』片段。Fab′通過切割Fab′2中連接重鏈的鉸鏈二硫鍵而在形式上衍生自Fab′2片段。除了抗體片段的二硫鍵連接以外的其它化學偶聯也是本領域已知的。Fab片段保留了親本抗體與其抗原結合的能力,潛在地具有更低的親和力。Fab′2片段能夠二價結合,而Fab和Fab』片段可以單價結合。通常,Fab片段缺乏恆定CH2和CH3域,即在其中將發生與Fc受體的相互作用的Fc部分。因此,Fab片段通常缺乏效應子功能。Fab片段可以通過本領域已知的方法通過抗體的酶切而產生,例如使用木瓜蛋白酶以獲得Fab或使用胃蛋白酶以獲得Fab′2,包括Fab、Fab′、Fab′2在內的Fab片段可以使用本領域技術人員公知的技術重組產生。
「Fv」片段是含有完全抗原識別和結合位點的抗體片段,且一般包含締合(其在性質上可以是共價的)的一個重鏈可變域和一個輕鏈可變域的二聚體,例如在單鏈可變域片段(scFv)中。以這種構型,每個可變域的三個高變區相互作用,以在VH-VL二聚體表面上限定抗原結合位點。六個高變區或其亞組共同地對抗體賦予抗原結合特異性。然而,即使僅包含三個抗原特異性高變區的單個可變域也可保留識別並結合抗原的能力,儘管其親和力通常低於完整的結合位點(Cai和Garen,Proc.Natl.Acad.Sci.USA(1996)93:6280-6285)。例如,僅具有重鏈可變域(VHH)的天然存在的駱駝科抗體可以結合抗原(Desmyter等人,J.Biol.Chem.,(2002)277:23645-23650;Bond等人,J.Mol.Biol.(2003)332:643-655)。
「單鏈Fv」或「scFv」抗體片段包含抗體的VH和VL域,其中這些結構域存在於單條多肽鏈中。通常,Fv多肽進一步在VH和VL域之間包含多肽接頭,該接頭使得scFv能夠形成用於抗原結合的所需結構。關於scFv的綜述,參見Pluckthun,1994,In:The Pharmacology of Monoclonal Antibodies,Vol.113,Rosenburg and Moore eds.Springer-Verlag,New York,pp.269-315。
術語「雙抗體(diabodies)」是指具有兩個抗原結合位點的小抗體片段,其中片段包含同一多肽鏈(VH和VL)中連接至輕鏈可變域(VL)的重鏈可變域(VH)。通過使用由於過短而無法允許在同一條鏈上的兩個可變域之間配對的接頭,使得這些可變域與另一條鏈的互補結構域配對,從而產生兩個抗原結合位點。雙抗體在例如EP 404,097、WO 93/11161以及Hollinger等人,1993,Proc.Natl.Acad.Sci.USA,90:6444-6448中更全面地描述。
表述「線性抗體」是指如在Zapata等人,1995,Protein Eng.8(10):1057-1062中描述的抗體。簡言之,這些抗體含有一對串聯的Fd區段(VH-CH1-VH-CH1),該區段與互補的輕鏈多肽一起形成一對抗原結合區。線性抗體可以是雙特異性的或單特異性的。
可採用常規的重組或蛋白質工程技術獲得抗體片段,並且可以以與完整抗體相同的方式,如本文所述,針對與TFPI(1-161)、全長TFPIα和TFPIβ或TFPI另一部分的結合篩選這些片段。
本發明的抗體片段可通過截短,例如通過從多肽的N-末端和/或C-末端去除一個或多個胺基酸來製備。也可以通過一個或多個內部缺失來生成片段。
本文中的術語「雙特異性抗體」是指具有兩個不同和/或獨特的抗原識別位點的抗體,這使抗體能夠結合兩種不同的抗原或同一抗原上的兩個不同表位。術語「多特異性抗體」是指具有結合兩種或更多種不同抗原或者同一抗原上兩個或更多個不同表位的能力的抗體。因此,多特異性抗體包括雙特異性抗體。
本發明的雙特異性抗體可以是IgG形式的全長雙特異性抗體。可通過融合兩個單獨的雜交瘤以形成產生包括雙特異性異二聚化抗體一部分的抗體混合物的雜交四源雜交瘤(quadroma),來產生模擬天然抗體的全長IgG形式的雙特異性抗體(Chelius D.等人;MAbs.2010年5-6月;2(3):309-319)。或者,可通過使用重組技術產生雙特異性異二聚化抗體。也可通過工程改造Fc區的二聚化界面以促進異二聚化來實現異二聚化。其中一個實例是所謂的杵臼(knob-in-hole)突變,其中將空間上大的側鏈(杵)引入一個Fc中,該Fc與相對Fc上空間上小的側鏈(臼)相匹配,從而創建促進異二聚化的空間互補。用於工程改造異二聚化Fc界面的其它方法是靜電互補,與非IgG異二聚化結構域融合,或利用人IgG4的天然Fab-臂交換現象來控制異二聚化。異二聚化的雙特異性抗體的實例在文獻中詳細描述,例如,(Klein C等人;MAbs.2012年11-12月;4(6):653-663)。必須特別注意異二聚體抗體中的輕鏈。可通過使用共同的輕鏈來實現LC與HC的正確配對。此外,LC/HC界面的工程化可用來促進異二聚化或如CrossMab中的輕鏈交叉工程化。來自含有適當突變的兩個單獨IgG的抗體在溫和還原條件下的體外重裝配還可用來產生雙特異性抗體(例如,Labrijn等人,PNAS,110,5145-5150(2013))。還報導了天然Fab-臂交換方法以確保正確的輕鏈配對。
還可將基於多特異性抗體的分子重組表達為組合了IgG的天然模塊的融合蛋白,以形成如文獻中所述的多特異性和多價抗體衍生物。融合抗體的實例是DVD-Ig、IgG-scFV、雙抗體、DART等(Kontermann,MAbs.2012年3-4月4(2):182-197)。可將特異性檢測或純化標籤、半衰期延長部分或其它組分併入融合蛋白中。也可將其它非IgG模塊併入融合蛋白中。不論LC配對方法如何,通常都將基於FC異二聚化的雙特異性全長抗體稱為不對稱IgG。
還可通過化學綴合或偶聯各個全長IgG或偶聯IgG的片段來產生基於多特異性抗體的分子,以形成如文獻中所述的多特異性和多價抗體衍生物。融合抗體的實例是化學偶聯的Fab』2、IgG二聚體等(Kontermann,MAbs.2012年3-4月4(2):182-197)。可將特異性檢測或純化標籤、半衰期延長分子或其它組分併入綴合蛋白中。還可將其它非IgG多肽併入融合蛋白中。在實施例中提供了這類雙特異性抗體的實例。
還可通過組合包括上述方法在內的重組和化學方法來產生多特異性分子。
本發明的雙特異性抗體可以是兩個抗原結合片段的化學綴合物,如兩個Fab片段(BiFab)或兩個scFv片段的綴合物。可通過本領域技術人員已知的方法來產生結合TFPI上的兩個獨特表位的雙特異性抗體或兩種單特異性抗體。例如,可通過兩個抗體片段,如兩個Fab或scFv片段的直接化學綴合或經由為正常功能提供所需柔性的接頭的化學綴合來製備雙特異性形式。一種用於偶聯Fab片段的具體方法是使用置於Fab片段適當位置的半胱氨酸殘基中的巰基官能團。
可通過兩個抗體或其片段(與TFPI的不同表位結合)的化學綴合來獲得本發明的雙特異性或雙功能性抗體,如J Immunol 1987;139:2367-2375或J Immunol 2001;166:1320-1326中所述,或如下所述:
mAb1(或片段)-mAb2(或片段)
mAb1(或片段)-接頭-mAb2(或片段)
連接可以是單個共價鍵(直接連接)或包含二價基團,該二價基團通常描述為:
其中X至少是但不限於一個選自碳、氧、硫、磷和氮的原子。*表示該二價基團的連接位置。術語「二價基團」是指具有兩個彼此獨立起作用的自由基中心的偶電子化學化合物。
在一個實施方案中,接頭是由不多於40個原子組成的鏈。
在一個實施方案中,在接頭中使用的化學部分包括具有以下結構的二價基團:
在一個實施方案中,接頭部分包含具有對稱結構的二價基團(同型雙功能化接頭)。
接頭還可以是具有與以上描述類似的結構的聚合物。
在一個實施方案中,在接頭中使用的化學部分包括聚合物:由通過共價化學鍵連接的兩個或更多個重複結構單元組成的大分子。這樣的聚合物可以是親水的。
術語親水的或「水溶性的」是指在水中具有某種可檢測程度的溶解度的部分。檢測和/或定量水溶解度的方法是本領域公知的。
根據本發明的示例性水溶性聚合物包括肽、糖、(聚)醚、(聚)胺、(聚)羧酸等。肽可具有混合序列或可由單種胺基酸組成,例如(聚)賴氨酸。一種示例性多糖是(聚)唾液酸。一種示例性(聚)醚是(聚)乙二醇。(聚)乙烯亞胺是一種示例性聚胺,而(聚)丙烯酸是一種典型的(聚)羧酸。
許多其它聚合物也適用於本發明。水溶性的聚合物主鏈在本發明中特別有用。合適的聚合物的實例包括但不限於,其它聚(亞烷基二醇)如聚(丙二醇)(「PPG」)、乙二醇和丙二醇的共聚物等,聚(氧乙基化多元醇),聚(乙烯吡咯烷酮),聚(羥丙基甲基丙烯醯胺),聚(α-羥酸),聚(乙烯醇),聚膦腈,聚噁唑啉,聚(N-烯丙醯嗎啉),如第5,629,384號美國專利中所述,其整體通過引用併入本文,以及它們的共聚物、三元共聚物及混合物。
聚合接頭優選是線性的。
儘管每一單獨聚合物鏈的分子量可以變化,但聚合物的平均分子量通常在約1000Da(1kDa)至約40,000Da(40kDa)的範圍內,諸如約1000Da至約12,000Da,如約2,000Da至約11,000Da,如約2000Da至約3,000Da;約3000Da至約4,000Da;約4000至約5,000Da;約5000至約6,000Da;約6,000至約7,000Da;約7,000至約8,000Da;約8,000至約9,000Da;約9,000至約10,000Da;或約10,000至約11,000Da。應當理解,這些大小表示估計值而非精確測量值。根據優選的實施方案,本發明的分子與親水性聚合物的異質群體綴合。
在一個具體實施方案中,在接頭中使用的化學部分包括聚乙二醇(PEG)。
本文中的術語「PEG」是指包含以下結構的二價基團:
其中n』是大於1的整數。
PEG通過環氧乙烷的聚合反應來製備,並且可在大範圍的分子量下商購獲得。根據本發明使用的PEG優選是線性的。
此外,「PEG」可指具有或不具有偶聯劑、偶聯或活化部分(例如,具有羧酸/活性酯、酮基、烷氧基胺、硫醇、三氟甲磺酸酯、三氟乙基磺酸酯(tresylate)、吖丙啶、環氧乙烷、炔、疊氮化物或馬來醯亞胺部分)的聚乙二醇化合物或其衍生物。本文提及的其它接頭也可以具有或不具有偶聯劑、偶聯或活化部分(例如,具有羧酸/活性酯、酮基、烷氧基胺、硫醇、三氟甲磺酸酯、三氟乙基磺酸酯、吖丙啶、環氧乙烷、炔、疊氮化物或馬來醯亞胺部分)
在一個具體實施方案中,根據本發明使用的PEG是單分散的。在另一具體實施方案中,根據本發明使用的PEG是多分散的。
多分散PEG由具有不同分子量的PEG分子組成。大小分布可通過其重均分子量(Mw)和其數均分子量(Mn)在統計學上表徵,其比值被稱為多分散性指數(Mw/Mn)(參見例如「Polymer Synthesis and Characterization」,J.A.Nairn,University of Utah,2003)。可通過質譜法來測量Mw和Mn。
多分散性指數可以是大於或等於1的數值,並且可由凝膠滲透色譜數據估算。當多分散性指數為1時,產物是單分散的,並因此由具有單一分子量的化合物組成。當多分散性指數大於1時,聚合物是多分散的,並且該多分散性指數表明具有不同分子量的聚合物的分布廣泛性。多分散性指數通常隨著PEG分子量的增加而增加。在具體實施方案中,根據本發明使用的PEG的多分散性指數:i)低於1.06,ii)低於1.05,iii)低於1.04,iv)低於1.03,或v)在1.02與1.03之間。
根據用於聚合過程的起始劑,可獲得不同形式的PEG。
在Advanced Drug Delivery Reviews,2002,54,459-476,Nature Reviews Drug Discovery,2003,2,214-221DOI:10.1038/nrd1033,Adv Polym Sci,2006,192,95-134,DOI 10.1007/12_022,Springer-Verlag,Berlin Heidelberg,2005,以及其中的參考文獻中描述了用於PEG取代基綴合的多種方法。或者,可通過使用酶促方法來進行親水聚合物取代基的綴合。這樣的方法例如是使用如WO2006134148中所述的轉穀氨醯胺酶。
為了實現聚合物分子與多肽的共價連接,以活化形式提供聚合物分子的端基,即具有反應性官能團。合適的活化聚合物分子可商購獲得,例如來自Aldrich Corporation,St.Louis,MO,USA,Rapp Polymere GmbH,Tübingen,Germany或來自PolyMASC Pharmaceuticals plc,UK。或者,可通過本領域已知的常規方法激活聚合物分子,例如,如WO 90/13540中所公開的。在第5,932,462和5,643,575號美國專利中公開了活化的PEG聚合物的具體實例。此外,下列公開文本公開了有用的聚合物分子和/或聚乙二醇化化學反應:WO2003/031464,WO2004/099231。
可通過使用任何常規方法來進行單克隆抗體或其片段與活化的聚合物分子的綴合,例如,如下列參考文獻(其也描述了用於聚合物分子活化的合適方法)所述:R.F.Taylor,(1991),″Protein immobilisation.Fundamental and applications″,Marcel Dekker,N.Y.;S.S.Wong,(1992),″Chemistry of Protein Conjugation and Crosslinking″,CRC Press,Boca Raton;G.T.Hermanson等人,(1993),″Immobilized Affinity Ligand Techniques″,Academic Press,N.Y.,Bioconjugate Techniques,第二版,Greg T.Hermanson,2008,Amsterdam,Elsevier。熟練的技術人員將意識到,待使用的活化方法和/或綴合化學反應取決於多肽的連接基團(其實例在上文進一步給出)以及聚合物的官能團(例如,為胺、羥基、羧基、醛、巰基、琥珀醯亞胺基、馬來醯亞胺、乙烯基碸或滷代乙酸酯)。聚乙二醇化可針對與多肽上可用的連接基團(即暴露在多肽表面的這類連接基團)綴合,或者可針對一個或多個特定連接基團,例如N-末端氨基或巰基。此外,可在一個步驟中或以逐步的方式實現綴合。
在另一實施方案中,用作接頭的化學部分是羥乙基澱粉。如本文中所用的術語「羥乙基澱粉」(HES/HAES)是指非離子澱粉衍生物。不同類型的羥乙基澱粉通常通過它們的平均分子量進行描述,通常約為130至200kDa。
在另一實施方案中,在接頭中使用的化學部分包括聚唾液酸。
在另一實施方案中,在接頭中使用的化學部分包括在例如Glycobiology(2011)21:1331-1340中描述的肝素前體(heparosan)聚合物。
在另一實施方案中,使用接頭中的化學部分使至少一種蛋白質與聚糖連接:與蛋白質連接的多糖或寡糖。
在另一實施方案中,使用接頭中的化學部分使至少一種蛋白質與O-連接的聚糖連接。
在另一實施方案中,使用接頭中的化學部分使至少一種蛋白質與N-連接的聚糖連接。
通過產生這些蛋白質的細胞使N-聚糖和O-聚糖均與蛋白質如抗體連接。當新生蛋白從核糖體轉移至內質網時,細胞N-糖基化機制識別胺基酸鏈中的N-糖基化信號(N-X-S/T基序)並使其糖基化(Kiely等人,J.Biol.Chem.1976,251:5490;Glabe等人,J.Biol.Chem.,1980,255,9236)。同樣,使O-聚糖連接至胺基酸鏈中的特異性O-糖基化位點,但是觸發O-糖基化的基序比N-糖基化信號更異質,並且我們預測胺基酸序列中O-糖基化位點的能力仍然不足(Julenius等人,Glycobiology,2005,15:153)。例如在WO0331464中描述了使多肽與各種聚合的側基綴合的方法。
在另一實施方案中,在接頭中使用的化學部分包括用來連接所述接頭與至少一種蛋白質的化學部分,具有以下二價基團的所選結構:
本發明的雙特異性抗體可以是人、人源化、嵌合或工程抗體。
如本文所用的,術語「人抗體」旨在包括具有可變區的抗體,在該可變區中,框架區的至少一部分和/或CDR區的至少一部分來源於人種系免疫球蛋白序列。(例如,人抗體可具有其中框架區和CDR區均來源於人種系免疫球蛋白序列的可變區。)此外,如果抗體含有恆定區,則該恆定區或其一部分也來源於人種系免疫球蛋白序列。本發明的人抗體可包括不是由人種系免疫球蛋白序列編碼的胺基酸殘基(例如,通過體外隨機或位點特異性突變或通過體內體細胞突變引入的突變)。
這樣的人雙特異性抗體可以來源於兩種人單克隆抗體。人單克隆抗體可以由雜交瘤產生,該雜交瘤包括與無限增殖化細胞融合的B細胞,該B細胞從轉基因非人類動物例如轉基因小鼠獲得,具有包含人免疫球蛋白重鏈和輕鏈基因區段的所有組成成分的基因組。
人抗體可以從基於人種系序列的選擇而建立的、用天然和合成序列多樣性進一步多樣化的序列文庫中分離。
人抗體可通過人淋巴細胞的體外免疫及隨後用EB病毒轉化該淋巴細胞來製備。
術語「人抗體衍生物」是指人抗體的任何修飾形式,如抗體與另一種物質或抗體的綴合物。
如本文所用的,術語「人源化抗體」是指含有來源於非人免疫球蛋白的序列元件(CDR區或其部分)的人/非人嵌合抗體。因此,人源化抗體是這樣的人免疫球蛋白(接受體抗體),其中至少來自該接受體的高變區的殘基被來自非人類物種如小鼠、大鼠、兔或非人靈長類動物的抗體(供體抗體)的高變區的、具有所需特異性、親和力、序列組成和功能性的殘基所替代。在一些情況下,人免疫球蛋白的框架(FR)殘基被相應的非人類殘基所替代。這種修飾的一個實例是一個或多個所謂的回覆突變的引入,該回復突變一般是來源於供體抗體的胺基酸殘基。抗體的人源化可採用本領域技術人員已知的重組技術進行(參見,例如,Antibody Engineering,Methods in Molecular Biology,vol.248,由Benny K.Lo編著)。對輕鏈和重鏈可變域均適合的人類接受體框架可通過例如序列或結構同源性來鑑定。或者,例如,可基於對結構、生物物理學和生物化學性質的了解使用固定的接受體框架。該接受體框架可以是種系衍生的或衍生自成熟的抗體序列。來自供體抗體的CDR區可以通過CDR移植進行轉移。可通過確定關鍵框架位置來進一步在例如親和力、功能性和生物物理學性質方面優化CDR移植的人源化抗體,在該關鍵框架位置處再次引入(回復突變)來自供體抗體的胺基酸殘基對人源化抗體的性質具有有利影響。除了來源於供體抗體的回覆突變外,還可通過在CDR或框架區中引入種系殘基、消除免疫原性表位、定點突變、親和力成熟等對人源化抗體進行工程化。
此外,人源化抗體可包含在接受體抗體或供體抗體中未發現的殘基。進行這些修飾以進一步改善抗體性能。通常,人源化抗體將包含至少一個-一般兩個-可變域,其中全部或基本上全部的CDR區對應於非人免疫球蛋白的CDR區,並且其中全部或基本上全部的FR殘基是人免疫球蛋白序列的FR殘基。人源化抗體也可任選地包含免疫球蛋白恆定區的至少一部分(Fc),一般是人免疫球蛋白的相應部分。
術語「人源化抗體衍生物」是指人源化抗體的任何修飾形式,如抗體與另一種化學劑或抗體或抗體片段或多肽的綴合物。
如本文所用的,術語「嵌合抗體」是指這樣的抗體,其輕鏈和重鏈基因已經從起源於不同物種的免疫球蛋白可變區和恆定區基因構建,一般是通過遺傳工程化構建。例如,來自小鼠單克隆抗體的基因的可變區段可連接到人恆定區。
抗體的片段可結晶區(「Fc區」/「Fc域」)是包含恆定CH2和CH3域的抗體C-末端區。Fc域可與被稱為Fc受體的細胞表面受體以及補體系統的一些蛋白質相互作用。Fc區使得抗體能夠與免疫系統相互作用。在本發明的一個方面,可對抗體進行工程化以在Fc區內包含修飾,一般用於改變其一種或多種功能性質,如血清半衰期、補體固定、Fc受體結合、蛋白質穩定性和/或抗原依賴性細胞毒性,或這些性質的缺乏,等等。此外,可對本發明的抗體進行化學修飾(例如,可將一個或多個化學部分連接至該抗體)或進行修飾以改變其糖基化,從而再次改變該抗體的一種或多種功能性質。IgG1抗體可攜帶修飾的Fc域,該Fc域包含以下突變中的一個或多個以及可能全部,這些突變將分別導致對某些Fc受體的親和力降低(L234A、L235E和G237A)以及C1q介導的補體固定減少(A330S和P331S)(殘基根據EU索引編號)。
本發明抗體的同種型可以是IgG,如IgG1,如IgG2,如IgG4。如果需要,抗體的類別可通過已知技術進行「轉換」。例如,最初作為IgM分子產生的抗體可類別轉換為IgG抗體。類別轉換技術還可用來將一個IgG亞類轉換成另一個亞類,例如:從IgG1轉換成IgG2或IgG4;從IgG2轉換成IgG1或IgG4;或從IgG4轉換成IgG1或IgG2。還可以進行通過來自不同IgG亞類的區域的組合而生成恆定區嵌合分子的抗體工程化。
在一個實施方案中,對CH1的鉸鏈區進行修飾,使得該鉸鏈區中的半胱氨酸殘基的數目改變,例如增加或減少。該方法在例如Bodmer等人的第5,677,425號美國專利中進一步描述。
可對恆定區進行修飾以使抗體穩定化,例如,降低二價抗體分離成兩個單價VH-VL片段的風險。例如,在lgG4恆定區中,殘基S228(根據EU編號索引,根據Kabat為S241)可突變成脯氨酸(P)殘基,以使鉸鏈處的重鏈間二硫鍵形成穩定化(參見,例如,Angal等人,Mol Immunol.1993;30:105-8)。
本發明的雙特異性抗體可包含mAb 2F3抗體、mAb 2F22抗體(如Fab 0295或Fab 0296)、mAb 2F45抗體或mAb 1F91抗體的抗原結合片段或其變體。
本發明的雙特異性抗體可進一步包含mAb 2021抗體的抗原結合片段或其變體,該抗體是在WO2010/072691中首次描述的人源化單克隆抗體,並且由此通過引用而併入。這類抗原結合片段或其變體的實例包括如本文描述的Fab 0088、Fab 0094或Fab 0313。衍生出mAb 2021的單克隆鼠抗體可以如WO2010/072691所述產生。
包含mAb 2F3、mAb 2F22、mAb 2F45或mAb 1F91的抗原結合片段或其變體和mAb 2021的抗原結合片段或其變體的上述雙特異性抗體中的任一種可以是IgG形式的全長雙特異性抗體或由抗體片段組成的雙特異性分子。抗體或其片段可按照其互補決定區(CDR)來定義。術語「互補決定區」或「高變區」在本文中使用時是指參與抗原結合的胺基酸殘基位於其中的抗體區域。高變區或CDR可被鑑定為在抗體可變域的胺基酸比對中具有最高可變性的區域。資料庫如Kabat資料庫可用於CDR鑑定,例如,CDR被定義為包含輕鏈可變域的胺基酸殘基24-34(L1)、50-56(L2)和89-97(L3)以及重鏈可變域的31-35(H1)、50-65(H2)和95-102(H3);(Kabat等人,Sequences of Proteins of Immunological Interest,1991)Sequences of Proteins of Immunological Interest,第五版,U.S.Department of Health and Human Services,NIH Publication No.91-3242)。或者,CDR可被定義為來自「高變環」的那些殘基(輕鏈可變域中的殘基26-33(L1)、50-52(L2)和91-96(L3)以及重鏈可變域中的26-32(H1)、53-55(H2)和96-101(H3);Chothia和Lesk,J.Mol.Biol 1987;196:901-917)。通常,該區域中胺基酸殘基的編號通過在Kabat等人(同上)中描述的方法進行。本文中的短語如「Kabat位置」、「Kabat殘基」和「根據Kabat」是指用於重鏈可變域或輕鏈可變域的這一編號體系。通過使用Kabat編號體系,肽的實際線性胺基酸序列可含有較少的或額外的胺基酸,這對應於可變域的框架(FR)或CDR的縮短或向其中的插入。例如,重鏈可變域可包含在CDR H2的殘基52後的胺基酸插入(根據Kabat的殘基52a、52b和52c)以及在重鏈FR殘基82後插入的殘基(例如,根據Kabat的殘基82a、82b和82c等)。可通過將抗體序列的同源性區域與「標準的」Kabat編號的序列進行比對,來確定給定抗體的殘基的Kabat編號。
術語「框架區」或「FR」殘基是指如本文定義的不在CDR內的那些VH或VL胺基酸殘基。
本發明雙特異性抗體的抗原識別位點可包含來自一種或多種本文公開的特異性抗體或抗體片段的CDR區,如來自SEQ ID NO:4至11、14至15、17至26內的CDR區,如根據本文公開的連續胺基酸編號或使用Kabat編號所定義的。
所述雙特異性抗體的第一抗原識別位點可具有重鏈,該重鏈包含:
●對應於SEQ ID NO:4的胺基酸31至35(SYGVH)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:4的胺基酸50至65(VIWRGGSTDFNAAFMS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:4的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸置換。
並且所述第一抗原識別位點可具有輕鏈,該輕鏈包含:
●對應於SEQ ID NO:5的胺基酸24至34(KASENVGAAVA)的CDR1序列,其中這些胺基酸殘基中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:5的胺基酸50至56(SASNRYT)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;如/或
●對應於SEQ ID NO:5的胺基酸89至96(QQYTNYPT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
所述雙特異性抗體的第一抗原識別位點可具有重鏈,該重鏈包含:
●對應於SEQ ID NO:6的胺基酸31至35(NYGVH)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:6的胺基酸50至65(VIWRGGSIDYNAAFMS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:6的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸置換。
並且所述第一抗原識別位點可具有輕鏈,該輕鏈包含:
●對應於SEQ ID NO:7的胺基酸24至34(KASQSVGPAVA)的CDR1序列,其中這些胺基酸殘基中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:7的胺基酸50至56(SASNRYT)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:7的胺基酸89至96(QQYTSYPT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
所述雙特異性抗體的第一抗原識別位點可具有重鏈,該重鏈包含:
●對應於SEQ ID NO:8的胺基酸31至35(GYGVH)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:8的胺基酸50至65(VIWRGGSIDYNAAFMS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:8的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸置換。
並且所述第一抗原識別位點可具有輕鏈,該輕鏈包含:
●對應於SEQ ID NO:9的胺基酸24至34(KASQNVGTAVA)的CDR1序列,其中這些胺基酸殘基中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:9的胺基酸50至56(SASNRYT)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:9的胺基酸89至96(QQYTSYPT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
所述雙特異性抗體的第一抗原識別位點可具有重鏈,該重鏈包含:
●對應於SEQ ID NO:10的胺基酸31至36(SDYAWN)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:10的胺基酸51至66(YISYSGSTSYNPSLKS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:10的胺基酸99至104(WAYDGP)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸置換。
並且所述第一抗原識別位點可具有輕鏈,該輕鏈包含:
●對應於SEQ ID NO:11的胺基酸24至33(RASSSVSHMH)的CDR1序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:11的胺基酸49至55(ATSNLAS)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:11的胺基酸88至96(QQWSSNPFT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
所述雙特異性抗體的第二抗原識別位點可具有重鏈,該重鏈包含:
●對應於SEQ ID NO:19的胺基酸31至35(NYAMS)的CDR1序列,其中這些胺基酸中的一個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:19的胺基酸50至66(TISRSGSYSYFPDSVQG)、SEQ ID NO:21的胺基酸50至66(TISRSGSYSYYPDSVKG)或SEQ ID NO:25的胺基酸50至66(TISRSGSYSYYADSVKG)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:19的胺基酸99至110(LGGYDEGDAMDS)的CDR3序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換。
並且所述第二抗原識別位點可具有輕鏈,該輕鏈包含:
●對應於SEQ ID NO:20的胺基酸24至39(KSSQSLLESDGKTYLN)的CDR1序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:20的胺基酸55至61(LVSILDS)的CDR2序列,其中這些胺基酸中的一個或兩個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:20的胺基酸94至102(LQATHFPQT)的CDR3序列,其中這些胺基酸中的一個或兩個可被不同的胺基酸置換。
所述胺基酸置換可以是如下描述的保守置換。
因此,TFPI(1-79)特異性抗體片段與其KPI-2結構域特異性抗體片段組合以形成尤其為融合形式、IgG片段形式、全長IgG形式或作為Fab-Fab綴合物(BiFab)的雙特異性抗體。
在一個這樣的實施方案中,攜帶mAb 1F91的VH和VL區(SEQ ID NO:10,11)的Fab片段與攜帶mAb 2021的VH、VL區(SEQ ID NO:19,20)的Fab片段組合。
在一個這樣的實施方案中,攜帶mAb 1F91的VH和VL區(SEQ ID NO:10,11)的Fab片段與Fab片段0094(SEQ ID NO:21,22)組合。
在一個這樣的實施方案中,攜帶mAb 1F91的VH和VL區(SEQ ID NO:10,11)的Fab片段與Fab片段0313(SEQ ID NO:25,26)組合。
在一個這樣的實施方案中,攜帶mAb 2F3的VH和VL區(SEQ ID NO:4,5)的Fab片段與攜帶mAb 2021的VH、VL區(SEQ ID NO:19,20)的Fab片段組合。
在一個這樣的實施方案中,攜帶mAb 2F3的VH和VL區(SEQ ID NO:4,5)的Fab片段與Fab片段0094(SEQ ID NO:21,22)組合。
在一個這樣的實施方案中,攜帶mAb 2F3的VH和VL區(SEQ ID NO:4,5)的Fab片段與Fab片段0313(SEQ ID NO:25,26)組合。
在一個這樣的實施方案中,攜帶mAb 2F22的VH和VL區(SEQ ID NO:6,7)的Fab片段與攜帶mAb 2021的VH、VL區(SEQ ID NO:19,20)的Fab片段組合。
在一個這樣的實施方案中,攜帶mAb 2F22的VH和VL區(SEQ ID NO:6,7)的Fab片段與Fab片段0094(SEQ ID NO:21,22)組合。
在一個這樣的實施方案中,攜帶mAb 2F22的VH和VL區(SEQ ID NO:6,7)的Fab片段與Fab片段0313(SEQ ID NO:25,26)組合。
在一個這樣的實施方案中,攜帶mAb 2F45的VH、VL區(SEQ ID NO:8,9)的Fab片段與mAb 2021的Fab片段(SEQ ID NO:19,20)組合。
在一個這樣的實施方案中,攜帶mAb 2F45的VH和VL區(SEQ ID NO:8,9)的Fab片段與Fab片段0094(SEQ ID NO:21,22)組合。
在一個這樣的實施方案中,攜帶mAb 2F45的VH和VL區(SEQ ID NO:8,9)的Fab片段與Fab片段0313(SEQ ID NO:25,26)組合。
根據本發明的一種Fab-Fab綴合物(BiFab)可以是與Fab 0295(SEQ ID NO:15,18)綴合的BiFab 9041(Fab 0088(SEQ ID NO:23,24)。
根據本發明的另一種Fab-Fab綴合物(BiFab)可以是BiFab 9042,即與Fab 0295(SEQ ID NO:15,18)綴合的Fab 0313(SEQ ID NO:25,26)。
在一個實施方案中,全長形式的雙特異性抗體(mAb 0421)由來自mAb 0310(SEQ ID NO:27,28)和mAb 0336(SEQ ID NO:15,29)的單獨臂組成。變異抗體可以包含1、2、3、4、5、可達10個或更多個相對於以上討論的特定序列和片段的胺基酸置換和/或缺失和/或插入。「缺失」變體可以包含單獨胺基酸的缺失,小組胺基酸如1、2、3、4或5個胺基酸的缺失,或更大胺基酸區域的缺失,諸如特定胺基酸結構域或其它特徵的缺失。「插入」變體可以包含單獨胺基酸的插入,小組胺基酸如1、2、3、4或5個胺基酸的插入,或更大胺基酸區域的插入,諸如特定胺基酸結構域或其它特徵的插入。「置換」變體優選地涉及用相同數目的胺基酸替換一個或多個胺基酸,以及進行保守胺基酸置換。例如,可以將胺基酸置換為具有類似性質的替代胺基酸,例如,另一個鹼性胺基酸、另一個酸性胺基酸、另一個中性胺基酸、另一個帶電荷的胺基酸、另一個親水胺基酸、另一個疏水胺基酸、另一個極性胺基酸、另一個芳族胺基酸或另一個脂族胺基酸。可以用來選擇合適的取代基的20種主要胺基酸的某些性質如下:
優選的「衍生物」或「變體」包括這樣的「衍生物」或「變體」,其中出現在序列中的胺基酸不是天然存在的胺基酸,而是它們的結構類似物。也可以衍生化或修飾,例如標記在序列中使用的胺基酸,只要抗體的功能不會顯著地受到不利影響即可。
置換可以是但不限於保守置換。
在抗體合成過程中,或通過生產後修飾,或當抗體為重組形式時,可以使用定點突變、隨機突變或核酸的酶切和/或連接的已知技術製備如上所述的衍生物和變體。
本發明還涉及編碼本發明抗體的多核苷酸。因此,本發明的多核苷酸可編碼如本文描述的任何抗體。術語「核酸分子」和「多核苷酸」在本文中可互換使用,並且指代任意長度的核苷酸的聚合形式,該核苷酸是脫氧核糖核苷酸或核糖核苷酸,或它們的類似物。多核苷酸的非限制性實例包括基因、基因片段、信使RNA(mRNA)、cDNA、重組多核苷酸、質粒、載體、任意序列的分離的DNA、任意序列的分離的RNA、核酸探針和引物。本發明的多核苷酸可以以分離的或純化的形式提供。「編碼」所選擇的多肽的核酸序列是這樣的核酸分子,當置於適當調節序列的控制下時,其在體內被轉錄(在DNA的情況下)和翻譯(在mRNA的情況下)成多肽。編碼序列的邊界由在5′(氨基)末端的起始密碼子和在3′(羧基)末端的翻譯終止密碼子決定。對於本發明而言,這樣的核酸序列可以包括但不限於來自病毒、原核或真核mRNA的cDNA,來自病毒或原核DNA或RNA的基因組序列,以及甚至合成的DNA序列。轉錄終止序列可以位於編碼序列的3′側。
在一個實施方案中,本發明的多核苷酸包含編碼如上所述的VH或VL胺基酸序列的序列。例如,本發明的多核苷酸可以編碼如上所述包含SEQ ID NO:4-11、14-15或17-26中任一個的序列的多肽。或者,合適的多核苷酸序列可以是這些具體多核苷酸序列之一的變體。例如,變體可以是任何上述核酸序列的置換、缺失或添加變體。相對於在序列表中給出的序列,變異多核苷酸可以包含1、2、3、4、5、可達10、可達20、可達30、可達40、可達50、可達75個或更多個核酸置換和/或缺失。
合適的變體可與編碼本發明多肽的多核苷酸至少70%同源,優選至少80%或90%同源,更優選至少95%、97%或99%同源。測量同源性的方法是本領域公知的,並且本領域技術人員應當理解,在本文中,以核酸同一性為基礎來計算同源性。這樣的同源性可存在於至少15個,優選至少30個,例如至少40、60、100、200個或更多個連續核苷酸的區域上。這樣的同源性可存在於未修飾的多核苷酸序列的整個長度上。
測量多核苷酸同源性或同一性的方法是本領域已知的。例如,UWGCG程序包提供了可用來計算同源性的BESTFIT程序(例如以其默認設置使用)(Devereux等人(1984)Nucleic Acids Research 12:387-395)。
也可使用PILEUP和BLAST算法來計算同源性或排列序列(通常使用其默認設置),例如,如Altschul S.F.(1993)J Mol Evol 36:290-300;Altschul,S.F.等人(1990)J Mol Biol 215:403-10中所述。
用於進行BLAST分析的軟體可通過國家生物技術信息中心公開獲得(http://www.ncbi.nlm.nih.gov/)。該算法涉及首先通過識別當與資料庫序列中相同長度的字詞比對時匹配或滿足一些正值閾值得分T的查詢序列中長度為W的短字詞來鑑別高評分序列對(HSP)。T被稱為鄰近字詞得分閾值(Altschul等人,同上)。這些初始鄰近字詞命中(hit)充當用於啟動搜索以查找包含它們的HSP的種子(seed)。字詞命中沿著每個序列在兩個方向上延伸,使得累積比對得分可以增加。當出現以下情況時,每個方向上字詞命中的延伸停止:由於一個或多個負分殘基比對的累積使累積比對得分變為零或以下;或到達任一序列的末端。BLAST算法參數W、T和X決定了比對的靈敏度和速度。BLAST程序使用字詞長度(W)為11、BLOSUM62評分矩陣(參見Henikoff和Henikoff(1992)Proc.Natl.Acad.Sci.USA 89:10915-10919)比對(B)為50、期望值(E)為10、M=5、N=4以及兩條鏈的比較作為默認設置。
BLAST算法進行兩個序列之間相似性的統計分析;參見例如,Karlin和Altschul(1993)Proc.Natl.Acad.Sci.USA 90:5873-5787。
本文中的術語「抗原」(Ag)是指用於對免疫活性脊椎動物進行免疫以產生識別該Ag的抗體(Ab)的實體。在本發明的上下文中,合適的抗原包括人TFPI(1-161)和全長人TFPIα。本文中,Ag的含義更加廣泛,並且通常意在包括被Ab特異性識別的靶分子,因此包括在用於產生Ab的免疫過程或其它過程例如噬菌體展示中使用的分子片段或模擬物。
如本文所用的術語「表位」在「抗原結合多肽」如抗體(Ab)與其相應抗原(Ag)之間的分子相互作用的背景下定義。通常,「表位」是指在Ag上的、與Ab特異性結合的區或區域,即與Ab物理接觸的區或區域。物理接觸可採用針對Ab和Ag分子中的原子的各種標準(例如,距離截止值為如如如或溶劑可及性)來定義。蛋白質表位可包含Ag中的、直接參與Ab結合的胺基酸殘基(也稱為表位的免疫顯性組分)以及其它不直接參與結合的胺基酸殘基,如被Ab有效阻斷的Ag的胺基酸殘基,即在Ab的「溶劑排斥表面(solvent-excluded surface)」和/或「足跡」內的胺基酸殘基。
除非另有說明,否則本文中的術語「表位」包括TFPI任何特定區域中的、特異性結合本發明單特異性或雙特異性TFPI抗體或另一種TFPI特異性藥劑的兩種類型的結合區。TFPI可包含許多不同的表位,其可包括但不限於(1)線性肽表位;(2)由在成熟TFPI構象中彼此靠近的一個或多個非連續胺基酸組成的構象表位;以及(3)全部或部分由共價連接至TFPI的分子結構如碳水化合物基團組成的翻譯後表位。
可採用多種實驗和計算表位作圖方法在不同的詳細度水平上描述和表徵給定抗體(Ab)/抗原(Ag)對的表位。該實驗方法包括突變、X射線晶體學、核磁共振(NMR)譜法、氫氘交換質譜法(HX-MS)以及各種競爭結合方法;本領域已知的方法。由於每一種方法均依賴於獨特的原理,因此對表位的描述與確定表位的方法緊密相關。因此,根據所採用的表位作圖方法,給定Ab/Ag對的表位可有不同地描述。
在其最詳細的水平上,針對Ag與Ab之間相互作用的表位可通過限定存在於Ag-Ab相互作用中的原子接觸的空間坐標,以及關於其對結合熱力學的相對貢獻的信息來描述。在詳細度較低的水平上,表位可通過限定Ag與Ab之間的原子接觸的空間坐標來表徵。在詳細度更低的水平上,表位可通過其包含的胺基酸殘基來表徵,該胺基酸殘基根據特定的標準,如在Ab:Ag複合物中的原子之間的距離或原子的溶劑可及性來定義。在詳細度進一步更低的水平上,表位可通過功能,例如通過與其它Ab的競爭結合來表徵。表位也可以更泛泛地定義為包含這樣的胺基酸殘基,另一種胺基酸對該胺基酸殘基的置換將會改變Ab與Ag之間的相互作用特徵。
在通過Ab例如Fab片段與其Ag之間複合物的空間坐標限定的X-射線衍生晶體結構的語境中,除非另有說明或與上下文矛盾,否則術語表位在此被具體定義為在距Ab中的重原子的距離內具有重原子(即,非氫原子)的TFPI殘基。
根據所使用的表位作圖方法,在不同的詳細度水平上獲得對表位的描述和定義,基於這一事實,可以在不同的詳細度水平上類似地進行在相同Ag上針對不同Ab的表位比較。
如果在胺基酸水平上描述的,例如由X射線結構確定的表位含有相同的一組胺基酸殘基,則這些表位被稱為是相同的。如果表位共有至少一個胺基酸,則這些表位被稱為是重疊的。如果表位沒有共有胺基酸殘基,則這些表位被稱為是不同的(獨特的)。
術語「互補位」的定義通過反轉視角,來源於以上對「表位」的定義。因此,術語「互補位」是指在Ab上的、與Ag特異性結合的區或區域,即與Ag物理接觸的區或區域。
在通過Ab如Fab片段與其Ag之間複合物的空間坐標限定的X-射線衍生晶體結構的語境中,除非另有說明或與上下文矛盾,否則術語互補位在此被具體定義為其特徵在於在距TFPI中的重原子的距離內具有重原子(即,非氫原子)的Ag殘基。
給定抗體(Ab)/抗原(Ag)對的表位和互補位可通過常規方法來鑑定。例如,表位的通常位置可通過評估抗體與不同片段或變異TFPI多肽結合的能力來確定。在TFPI內的、與抗體(表位)接觸的具體胺基酸和在抗體中的、與TFPI(互補位)接觸的具體胺基酸也可使用常規方法來確定。例如,抗體和靶分子可以組合,並且可以使Ab:Ag複合物結晶。可以確定該複合物的晶體結構,並用其鑑定抗體與其靶標之間相互作用的特異性位點。
與相同抗原結合的抗體可就其同時與其共同抗原結合的能力進行表徵,並且可以經歷「競爭結合」/「分箱(binning)」。在該語境下,術語「分箱」是指對與相同抗原結合的抗體進行分組的方法。抗體的「分箱」可以基於在以標準技術為基礎的試驗中兩種抗體與其共同抗原的競爭結合,該標準技術例如是表面等離子體共振(SPR)、ELISA或流式細胞術。
使用參考抗體確定抗體的「箱元」。如果第二抗體不能與參考抗體同時與抗原結合,則稱第二抗體與參考抗體屬於同一「箱元」。在這種情況下,參考抗體和第二抗體競爭性地結合抗原的相同部分,因此被劃歸為「競爭抗體」。如果第二抗體能夠與參考抗體同時與抗原結合,則稱第二抗體屬於不同的「箱元」。在這種情況下,參考抗體和第二抗體不會競爭性地結合抗原的相同部分,因此被劃歸為「非競爭抗體」。
抗體「分箱」不提供關於表位的直接信息。競爭抗體,即屬於同一「箱元」的抗體,可具有相同的表位、重疊的表位或甚至不同的表位。如果與其在抗原上的表位結合的參考抗體佔據了第二抗體與其在抗原上的表位接觸所需的空間(「位阻」),則是後一種情況。非競爭抗體通常具有不同的表位。
術語「結合親和力」在本文中用作兩種分子(例如抗體或其片段和抗原)之間的非共價相互作用的強度的度量。術語「結合親和力」在本文中用於描述單價和雙價相互作用(內在活性)兩者。
通過測定平衡解離常數(KD),可以定量兩種分子(例如抗體或其片段和抗原)之間通過單價相互作用的結合親和力。反過來,通過測量複合物形成和解離的動力學,例如通過SPR方法,可以測定KD。與單價複合物的結合和解離相對應的速率常數分別被稱為結合速率常數ka(或k結合)和解離速率常數kd(或k解離)。通過方程式KD=kd/ka,將KD與ka和kd相關聯。
按照上面的定義,通過比較單個抗體/抗原複合物的KD值,可以比較與不同分子相互作用有關的結合親和力,例如比較不同抗體對給定抗原的結合親和力。
根據本發明的抗體可以能夠與另一種分子如天然存在的配體或受體或另一種抗體競爭結合TFPI,從而影響與這些相互作用相關的功能。抗體與天然配體/受體競爭的能力可以通過測量對TFPI抑制的表觀KI的影響的各種活性測定來評估。然後可以由表觀KI值推斷KD值。一般來說,抗體對於靶標(TFPI)的感興趣的KD值將為其它TFPI配體的KD的1/2,優選1/5,更優選1/10。更優選地,KD將為1/50,如1/100或1/200;甚至更優選1/500,如1/1000或1/10,000。
可以通過眾所周知的方法直接測定該解離常數的值。用來評價諸如抗體等配體與靶標的結合能力的標準試驗是本領域已知的,並且包括,例如,ELISA、Western印跡法、RIA、等溫滴定量熱法和流式細胞術分析。也可以通過本領域已知的標準試驗,如SPR,來評估抗體的結合動力學和結合親和力。
可以進行競爭結合試驗,其中將抗體與靶標的結合和該靶標的另一種配體如另一種抗體對該靶標的結合進行比較。
如本文公開的雙特異性抗體可以以比其各自「臂」的組合親和力更高的親和力和/或親合力結合全長TFPI或TFPI(1-161)。
如本文公開的雙特異性抗體可以以比其結合如本文公開的TFPI(1-96)、TFPI(1-76)、TFPI(26-76)、TFPI(77-161)、TFPI(97-147)和/或其它任何TFPI片段更高的親和力和/或親合力結合全長TFPI。
如本文公開的雙特異性抗體對全長TFPI或TFPI(1-161)可具有1×10-11M或更低,或1×10-12M或更低,或1×10-13M或更低,或1×10-14M或更低,或1×10-15M或更低的KD。
本發明雙特異性抗體的任何「臂」對其靶標可具有1×10-5M或更低,1×10-6M或更低,1×10-7M或更低,1×10-8M或更低,或1×10-9M或更低,或1×10-10M或更低,1×10-11M或更低,或1×10-12M或更低,或1×10-13M或更低,或1×10-14M或更低,或1×10-15M或更低的KD。本發明抗體的KD可以低於0.8nM,如低於0.7nM,如低於0.6nM,如低於0.5nM,如低於0.4nM,如低於0.3nM,如低於0.2nM,如低於0.1nM,如低於0.05nM,如低於0.025nM,如低於0.015nM,如介於0.015nM與0nM之間。
在一個方面,如本文公開的兩個(全長)抗體或Fab片段的雙特異性綴合物包含TFPI(1-79)結合子(binder)和TFPI KPI-2結合子,其中TFPI(1-79)結合子的KD為2×10-9M或更低,3×10-9M或更低,4×10-9M或更低,5×10-9M或更低,6×10-9M或更低,7×10-9M或更低,8×10-9M或更低,9×10-9M或更低,1×10-8M或更低,2×10-8M或更低,3×10-8M或更低,4×10-8M或更低,5×10-8M或更低,6×10-8M或更低,7×10-8M或更低,8×10-8M或更低,9×10-8M或更低,1×10-7M或更低,2×10-7M或更低,3×10-7M或更低,4×10-7M或更低,5×10-7M或更低,6×10-7M或更低,7×10-7M或更低,8×10-7M或更低,9×10-7M或更低,1×10-6M或更低,2×10-6M或更低,3×10-6M或更低,4×10-6M或更低,5×10-6M或更低,6×10-6M或更低,7×10-6M或更低,8×10-6M或更低,9×10-6M或更低,1×10-5M或更低,2×10-5M或更低,3×10-5M或更低,4×10-5M或更低,5×10-5M或更低,6×10-5M或更低,7×10-5M或更低,8×10-5M或更低,9×10-5M或更低。
在一個方面,TFPI(1-79)結合子的KD在5×10-7M至1×10-5M的範圍內。在一個這樣的方面,TFPI(1-79)結合子的KD在1×10-7M至1×10-5M的範圍內。在一個這樣的方面,TFPI(1-79)結合子的KD在1×10-7M至1×10-6M的範圍內。
在一個方面,TFPI(1-79)結合子的KD在5×10-7M至1×10-7M的範圍內。
在一個方面,TFPI(1-79)結合子的KD在1×10-7M至5×10-5M的範圍內。
在一個方面,TFPI(1-79)結合子是包含一個或多個置換的mAb2F22、Fab 0295或Fab 0296的變體。
在一個方面,KPI-2結合子是包含一個或多個置換的mAb 2021、mAb 0310、Fab 0088或Fab 0313的變體。
在一個方面,如本文公開的兩個(全長)抗體或Fab片段的雙特異性綴合物包含TFPI(1-79)結合子和TFPI KPI-2結合子,其中KPI-2結合子的KD為2×10-9M或更低,3×10-9M或更低,4×10-9M或更低,5×10-9M或更低,6×10-9M或更低,7×10-9M或更低,8×10-9M或更低,9×10-9M或更低,1×10-8M或更低,2×10-8M或更低,3×10-8M或更低,4×10-8M或更低,5×10-8M或更低,6×10-8M或更低,7×10-8M或更低,8×10-8M或更低,9×10-8M或更低,1×10-7M或更低,2×10-7M或更低,3×10-7M或更低,4×10-7M或更低,5×10-7M或更低,6×10-7M或更低,7×10-7M或更低,8×10-7M或更低,9×10-7M或更低,1×10-6M或更低,2×10-6M或更低,3×10-6M或更低,4×10-6M或更低,5×10-6M或更低,6×10-6M或更低,7×10-6M或更低,8×10-6M或更低,9×10-6M或更低,1×10-5M或更低,2×10-5M或更低,3×10-5M或更低,4×10-5M或更低,5×10-5M或更低,6×10-5M或更低,7×10-5M或更低,6×10-5M或更低,5×10-5M或更低,4×10-5M或更低,3×10-5M或更低,2×10-5M或更低,1×10-5M或更低。
在一個方面,KPI-2結合子的KD在5×10-7M至1×10-5M的範圍內。在一個這樣的方面,KPI-2結合子的KD在1×10-7M至1×10-5M的範圍內。在一個這樣的方面,KPI-2結合子的KD在1×10-7M至1×10-6M的範圍內。
在一個方面,KPI-2結合子的KD在5×10-7M至1×10-7M的範圍內。
在一個方面,KPI-2結合子的KD在1×10-7M至5×10-5M的範圍內。
在一個方面,如本文公開的雙特異性抗體包含KD在1×10-9M至1×10-6M範圍內的TFPI(1-79)結合子和KD在1×10-14M至1×10-10M範圍內的KPI-2結合子。在一個這樣的方面,TFPI(1-79)結合子KD在1×10-8M至1×10-6M的範圍內且KPI-2結合子KD在1×10-14M至1×10-11M的範圍內。
在一個方面,如本文公開的雙特異性抗體包含KD在1×10-14M至1×10-10M範圍內的TFPI(1-79)結合子和KD在1×10-12M至1×10-11M範圍內的KPI-2結合子。
在一個方面,如本文公開的雙特異性抗體包含KD在1×10-12M至1×10-10M範圍內的TFPI(1-79)結合子和KD在1×10-12M至1×10-8M範圍內的KPI-2結合子。在一個這樣的方面,TFPI(1-79)結合子的KD在1×10-11M至5×10-10M的範圍內且KPI-2結合子的KD在1×10-10M至1×10-8M的範圍內。
在一個方面,TFPI(1-79)結合子是mAb 0336且KPI-2結合子是mAb 0310。
在一個方面,KPI-2結合子是Fab 0088。在一個替代方面,KPI-2結合子是Fab 0094。在另一替代方面,KPI-2結合子是Fab 0313。
在一個方面,Fab 0094、Fab 0313、Fab 0094或Fab 0088與KPI-1結合子組合,該KPI-1結合子是包含一個或多個置換的Fab 0295的變體。
在一個方面,通過將兩個分別與TFPI(1-79)和TFPI KPI-2結合的抗體片段組合成雙特異性抗體而獲得對凝血酶生成的協同效應。
在一個方面,通過將兩個分別與TFPI(1-79)和TFPI KPI-2結合的抗體片段組合成雙特異性Fab(BiFab)而獲得對凝血酶生成的協同效應。
在一個方面,甚至在雙特異性抗體或Fab的一個臂對其靶標的親和力降低的情況下,也可以存在以上提到的協同效應。
在一個方面,本發明的雙特異性抗體可與FXa、TF/FVIIa和/或TF/FVIIa/FXa競爭結合TFPI。
在另一個方面,本發明提供了包含本發明分子,如本文所述雙特異性抗體的組合物和製劑。例如,本發明提供了包含與藥學上可接受的載體一起配製的一種或多種本發明雙特異性TFPI抗體的藥物組合物。
相應地,本發明的一個目的在於提供一種藥物製劑,其包含以0.25mg/ml至250mg/ml的濃度存在的這樣的雙特異性TFPI抗體,並且其中所述製劑具有2.0至10.0的pH。該製劑可以進一步包含緩衝體系、防腐劑、張度劑、螯合劑、穩定劑或表面活性劑及其各種組合中的一種或多種。防腐劑、等滲劑、螯合劑、穩定劑和表面活性劑在藥物組合物中的使用是技術人員公知的。可參考Remington:The Science and Practice of Pharmacy,第19版,1995。
在一個實施方案中,所述藥物製劑是水性製劑。這樣的製劑一般是溶液或懸浮液,但也可以包括膠體、分散體、乳液和多相物質。術語「水性製劑」被定義為包含至少50%w/w水的製劑。同樣,術語「水性溶液」被定義為包含至少50%w/w水的溶液,而術語「水性懸浮液」被定義為包含至少50%w/w水的懸浮液。
在另一個實施方案中,所述藥物製劑是冷凍乾燥的製劑,在使用前醫師或患者向其中添加溶劑和/或稀釋劑。
在進一步的方面,所述藥物製劑包含這樣的抗體的水性溶液和緩衝液,其中該抗體以1mg/ml或以上的濃度存在,並且其中所述製劑具有約2.0至約10.0的pH。
雙特異性抗體或包含該抗體的藥物製劑可以用來治療具有凝血障礙的受試者。
如本文所用的,術語「受試者」包括任何人類患者,或非人類脊椎動物。
如本文所用的,術語「凝血障礙」是指增加的出血傾向,這可由正常凝血級聯的任意促凝血組分的任何定性或定量缺乏或纖維蛋白溶解的任何上調引起。這類凝血障礙可能是先天性的和/或獲得性的和/或醫源性的,並且由本領域技術人員確定。
先天性低凝血障礙(hypocoagulopathies)的非限制性實例為A型血友病、B型血友病、因子VII缺乏、因子X缺乏、因子XI缺乏、馮威勒布蘭德氏病和血小板減少症如Glanzmann氏血小板機能不全(thombasthenia)和貝-索(Bernard-Soulier)症候群。所述A型或B型血友病可能是重度的、中度的或輕度的。血友病的臨床嚴重程度根據血液中的FIX/FVIII功能單元的濃度來確定,並被分類為輕度的、中度的或重度的。重度血友病被定義為凝血因子水平<0.01U/ml,對應於小於正常水平的1%,而中度和輕度的患者分別具有1-5%和>5%的水平。具有「抑制物」(即抗因子VIII的同種抗體)的A型血友病和具有「抑制物」(即抗因子IX的同種抗體)的B型血友病是部分先天性和部分獲得性的凝血障礙的非限制性實例。
獲得性凝血障礙的非限制性實例是由維生素K缺乏引起的絲氨酸蛋白酶缺乏;這樣的維生素K缺乏可能由維生素K拮抗劑如華法林的施用而引起。獲得性凝血障礙也可能在大面積創傷後發生。在這種也稱為「血性惡性循環」的情況下,其特徵為血液稀釋(稀釋性血小板減少症和凝血因子的稀釋)、低體溫、凝血因子的消耗和代謝紊亂(酸中毒)。補液療法和增加的纖維蛋白溶解可加劇這種情況。所述出血可能來自身體的任何部分。
醫源性凝血障礙的一個非限制性實例是可能為了治療血栓栓塞性疾病而開出的抗凝血藥物如肝素、阿司匹林、華法林及其它血小板聚集抑制劑的過高劑量。醫源性凝血障礙的第二個非限制性實例是由過度的和/或不當的補液療法誘發的凝血障礙,諸如可能由輸血誘發的凝血障礙。
在本發明的一個實施方案中,出血與A型或B型血友病有關。在另一個實施方案中,出血與具有獲得性抑制物的A型或B型血友病有關。在另一個實施方案中,出血與血小板減少症有關。在另一個實施方案中,出血與馮威勒布蘭德氏病有關。在另一個實施方案中,出血與嚴重組織損傷有關。在另一個實施方案中,出血與嚴重創傷有關。在另一個實施方案中,出血與手術有關。在另一個實施方案中,出血與出血性胃炎和/或腸炎有關。在另一個實施方案中,出血是血崩,諸如在胎盤早剝中。在另一個實施方案中,出血發生在機械性止血可能性受限的器官中,諸如顱內、耳內或眼內。在另一個實施方案中,出血與抗凝治療有關。
如本文所用的,術語「治療」是指對任何有需要的人類或其它脊椎動物受試者的藥物療法。所述受試者預期已經經歷了由執業醫師或執業獸醫進行的體格檢查,該醫師或獸醫已經給出了初步或明確的診斷,該診斷指明所述特定治療的使用對所述人類或其它脊椎動物的健康是有益的。根據受試者健康的現狀,所述治療的時間安排和目的可隨個體的不同而不同。因此,所述治療可以是預防性的、姑息性的、對症的和/或治癒性的。就本發明而言,預防性的、姑息性的、對症的和/或治癒性的治療可代表本發明的單獨的方面。
本發明的抗體可以腸胃外施用,如靜脈內,如肌肉內,如皮下施用。皮下施用是優選的。或者,本發明的抗體可通過非腸胃外途徑如經口或局部施用。本發明的抗體可以預防性施用。本發明的抗體可治療性地施用(根據需求)。
在一個實施方案中,本發明的雙特異性抗體用來在體外測定TFPIα水平。
以下是本發明的非限制性實施方案:
實施方案1:一種能夠特異性結合人TFPI(SEQ ID NO:1)的位置1至161內的第一表位和第二表位的雙特異性抗體或BiFab。
實施方案2:根據實施方案1的雙特異性抗體,其中所述第一表位在人TFPI的位置1-76內且所述第二表位在位置77-161內。
實施方案3:根據實施方案1的雙特異性抗體,其中所述第一表位在人TFPI的位置1-96內且所述第二表位在位置97-161內。
實施方案4:根據實施方案2-3中任一項的雙特異性抗體,其中所述第一表位在人TFPI的Kunitz型抑制劑1結構域(殘基26-76)內。
實施方案5:根據實施方案2-3中任一項的雙特異性抗體,其中所述第二表位在人TFPI的Kunitz型抑制劑2結構域(殘基97-147)內。
實施方案6:根據實施方案5的雙特異性抗體,其中所述KPI-1表位包含SEQ ID NO:1的胺基酸殘基Arg 41、Arg 65和/或Glu 67。
實施方案7:根據實施方案4或6中任一項的雙特異性抗體,其中所述KPI-1表位包含SEQ ID NO:1的胺基酸殘基Leu 16、Pro 17、Leu 19、Lys 20、Leu 21、Met 22、Phe 25、Cys 35、Ala 37、Met 39、Arg 41、Tyr 56、Gly 57、Gly 58、Cys 59、Glu 60、Gly 61、Asn 62、Gln 63、Arg 65、Phe 66、Glu 67、Glu 71和Met 75。
實施方案8:根據實施方案5的雙特異性抗體,其中所述第二表位包含SEQ ID NO:1的胺基酸殘基Arg 107。
實施方案9:根據實施方案5的雙特異性抗體,其中所述第二表位包含SEQ ID NO:1的胺基酸殘基Glu 100、Glu 101、Asp 102、Pro 103、Arg 107、Tyr 109、Thr 111、Tyr 113、Phe 114、Asn 116、Gln 118、Gln 121、Cys 122、Glu 123、Arg 124、Phe 125、Lys 126和Leu 140。
實施方案10:根據實施方案1的雙特異性抗體,其中第一抗原識別位點的重鏈包含:
●對應於SEQ ID NO:4的胺基酸31至35(SYGVH)、SEQ ID NO:6的胺基酸31至35(NYGVH)或SEQ ID NO:8的胺基酸31至35(GYGVH)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:4的胺基酸50至65(VIWRGGSTDFNAAFMS)、SEQ ID NO:6的胺基酸50至65(VIWRGGSIDYNAAFMS)或SEQ ID NO:8的胺基酸50至65(VIWRGGSIDYNAAFMS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:4的胺基酸98至110(NSHGNYVGYAMDY)、SEQ ID NO:6的胺基酸98至110或SEQ ID NO:8的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸置換;
並且其中所述第一抗原識別位點的輕鏈包含:
●對應於SEQ ID NO:5的胺基酸24至34(KASENVGAAVA)、SEQ ID NO:7的胺基酸24至34(KASQSVGPAVA)或SEQ ID NO:9的胺基酸24至34(KASQNVGTAVA)的CDR1序列,其中這些胺基酸殘基中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:5的胺基酸50至56(SASNRYT)、SEQ ID NO:7的胺基酸50至56或SEQ ID NO:9的胺基酸50至56(SASNRYT)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:5的胺基酸89至96(QQYTNYPT)、SEQ ID NO:7的胺基酸89至96(QQYTSYPT)或SEQ ID NO:9的胺基酸89至96(QQYTSYPT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
實施方案11:根據實施方案1的雙特異性抗體,其中第一抗原識別位點的重鏈包含:
●對應於SEQ ID NO:4的胺基酸31至35(SYGVH)、SEQ ID NO:6的胺基酸31至35(NYGVH)或SEQ ID NO:8的胺基酸31至35(GYGVH)的CDR1序列;和
●對應於SEQ ID NO:4的胺基酸50至65(VIWRGGSTDFNAAFMS)、SEQ ID NO:6的胺基酸50至65(VIWRGGSIDYNAAFMS)或SEQ ID NO:8的胺基酸50至65(VIWRGGSIDYNAAFMS)的CDR2序列;和
●對應於SEQ ID NO:4的胺基酸98至110、SEQ ID NO:6的胺基酸98至110或SEQ ID NO:8的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列;
並且其中所述第一抗原識別位點的輕鏈包含:
●對應於SEQ ID NO:5的胺基酸24至34(KASENVGAAVA)、SEQ ID NO:7的胺基酸24至34(KASQSVGPAVA)或SEQ ID NO:9的胺基酸24至34(KASQNVGTAVA)的CDR1序列;和
●對應於SEQ ID NO:5的胺基酸50至56、SEQ ID NO:7的胺基酸50至56或SEQ ID NO:9的胺基酸50至56(SASNRYT)的CDR2序列;和
●對應於SEQ ID NO:5的胺基酸89至96(QQYTNYPT)、SEQ ID NO:7的胺基酸89至96(QQYTSYPT)或SEQ ID NO:9的胺基酸89至96(QQYTSYPT)的CDR3序列。
實施方案12:根據實施方案1的雙特異性抗體,其中第一抗原識別位點的重鏈包含:
●對應於SEQ ID NO:6的胺基酸31至35(NYGVH)的CDR1序列;和
●對應於SEQ ID NO:6的胺基酸50至65(VIWRGGSIDYNAAFMS)的CDR2序列;和
●對應於SEQ ID NO:6的胺基酸98至110(NSHGNYVGYAMDY)的CDR3序列;
並且其中所述第一抗原識別位點的輕鏈包含:
●對應於SEQ ID NO:7的胺基酸24至34(KASQSVGPAVA)的CDR1序列;和
●對應於SEQ ID NO:7的胺基酸50至56(SASNRYT)的CDR2序列;和
●對應於SEQ ID NO:7的胺基酸89至96(QQYTSYPT)的CDR3序列。
實施方案13:根據實施方案1的雙特異性抗體,其中第一抗原識別位點的重鏈包含:
●對應於SEQ ID NO:10的胺基酸31至36(SDYAWN)的CDR1序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:10的胺基酸51至66(YISYSGSTSYNPSLKS)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:10的胺基酸99至104(WAYDGP)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸置換,
並且其中所述第一抗原識別位點的輕鏈包含:
●對應於SEQ ID NO:11的胺基酸24至33(RASSSVSHMH)的CDR1序列,其中這些胺基酸殘基中的一個或兩個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:11的胺基酸49至55(ATSNLAS)的CDR2序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換;和/或
●對應於SEQ ID NO:11的胺基酸88至96(QQWSSNPFT)的CDR3序列,其中這些胺基酸殘基中的一個可被不同的胺基酸殘基置換。
實施方案14:根據實施方案10-13中任一項的雙特異性抗體,其中第二抗原識別位點的重鏈包含:
●對應於SEQ ID NO:19的胺基酸31至35(NYAMS)的CDR1序列,其中這些胺基酸中的一個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:19的胺基酸50至66(TISRSGSYSYFPDSVQG)、SEQ ID NO:21的胺基酸50至66(TISRSGSYSYYPDSVKG)或SEQ ID NO:25的胺基酸50至66(TISRSGSYSYYADSVKG)的CDR2序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:19的胺基酸99至110(LGGYDEGDAMDS)的CDR3序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換。
並且其中所述第二抗原識別位點的輕鏈包含:
●對應於SEQ ID NO:20的胺基酸24至39(KSSQSLLESDGKTYLN)的CDR1序列,其中這些胺基酸中的一個、兩個或三個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:20的胺基酸55至61(LVSILDS)的CDR2序列,其中這些胺基酸中的一個或兩個可被不同的胺基酸置換;和/或
●對應於SEQ ID NO:20的胺基酸94至102(LQATHFPQT)的CDR3序列,其中這些胺基酸中的一個或兩個可被不同的胺基酸置換。
實施方案15:根據實施方案10-14中任一項的雙特異性抗體,其中所述置換是保守置換。
實施方案16:根據前述實施方案中任一項的雙特異性抗體,其選自全長雙特異性抗體和兩個抗原結合片段的化學綴合物。
實施方案17:根據實施方案16的雙特異性抗體,其為兩個Fab片段的綴合物。
實施方案18:根據實施方案1-17中任一項的雙特異性抗體,其為人源化抗體或人抗體。
實施方案19:根據實施方案1-18中任一項的雙特異性抗體,其對全長TFPI或TFPI(1-161)具有1×10-11M或更低,或1×10-12M或更低,或1×10-13M或更低,或1×10-14M或更低,或1×10-15M或更低的結合親和力。
實施方案20:一種藥物組合物,其包含根據實施方案1至18中任一項的雙特異性抗體和藥學上可接受的載體。
實施方案21:根據實施方案1至19中任一項的雙特異性抗體或根據實施方案20的藥物組合物,其用作藥物。
實施方案22:根據實施方案1至19中任一項的雙特異性抗體或根據實施方案20的藥物組合物,其用於治療凝血障礙。
實施方案23:用於根據實施方案22使用的雙特異性抗體,其中所述受試者具有先天性、獲得性和/或醫源性凝血障礙,如具有或不具有抑制物的A型血友病,或具有或不具有抑制物的B型血友病。
實施方案24:一種治療具有凝血障礙的受試者的方法,其包括向所述受試者施用根據實施方案1至19中任一項的雙特異性抗體。
實施方案25:根據實施方案1至19中任一項的雙特異性抗體在製備用於治療凝血障礙的藥物中的用途。
實施方案26:根據實施方案25的用途,其中所述凝血障礙是先天性、獲得性和/或醫源性凝血障礙。
實施方案27:根據實施方案26的用途,其中所述凝血障礙是具有或不具有抑制物的A型血友病,或具有或不具有抑制物的B型血友病。
本發明通過以下實施例進一步說明,這些實施例不應理解為進一步限制。所有附圖和整個本申請中引用的所有參考文獻、專利和公開的專利申請的內容均明確地通過引用併入本文。
實施例
實施例1:免疫、融合和篩選
用人TFPIα(SEQ ID NO:1)或TFPI(1-161)(SEQ ID NO:2)免疫RBF小鼠。向小鼠皮下注射:將20μg人TFPI與完全弗氏佐劑混合以供第一次注射。對於隨後的免疫,使用不完全弗氏佐劑與相同濃度的抗原。在最後免疫後10天,使用ELISA針對人TFPI特異性抗體篩選來自小鼠的眼部血液。通過靜脈內注射用10μg人TFPIα或TFPI(1-161)加強免疫具有陽性血清滴度的小鼠,並在三天後處死。在無菌條件下取出脾臟,並將其分散成單細胞懸浮液。藉助於PEG方法或通過電融合進行脾細胞與骨髓瘤細胞的融合。通過有限稀釋將所得雜交瘤細胞克隆至微量滴定板中。首先通過ELISA針對能夠與全長TFPIα或TFPI(1-161)結合的抗體的表達篩選來自單個雜交瘤的上清液。為了鑑別產生對TFPI(1-79)為特異性的抗體的雜交瘤,針對與TFPI KPI-2片段(由SEQ ID NO:1的胺基酸殘基97-147表示)的結合,反向篩選與全長TFPIα或TFPI(1-161)的結合呈陽性的雜交瘤。分離並擴增與TFPI(1-161)的結合呈陽性且與TFPI KPI-2片段的結合呈陰性的雜交瘤,以產生抗體。
通過標準蛋白A親和色譜法從上清液中純化抗體,並用其確定與人TFPIα和TFPI(1-79)的結合和親和力,以及血漿中的TFPI中和活性(TGT試驗)。通過有限稀釋對產生感興趣抗體的雜交瘤(即,對TFPI(1-79)為特異性的那些)進行亞克隆,並且針對來自亞克隆雜交瘤的物質驗證原始抗體譜。來自亞克隆雜交瘤的細胞用於分離RNA以及隨後的抗體克隆和序列鑑定。
實施例2:小鼠抗人TFPI(1-79)特異性抗體的克隆和測序
本實施例描述了TFPI抗體mAb 2F3、mAb 2F22、mAb 2F45、mAb 1F56和mAb 1F91的鼠重鏈和輕鏈序列的克隆和測序。使用來自Qiagen的RNeasy-Mini試劑盒從雜交瘤細胞中提取總RNA,並將其用作cDNA合成的模板。使用來自Clontech的SMARTTM RACE cDNA擴增試劑盒在5′-RACE反應中合成cDNA。使用Phusion Hot Start聚合酶(Finnzymes)和作為正向引物包含在SMARTTM RACE試劑盒中的通用引物混合物(UPM),通過PCR進行隨後的HC和LC序列的靶擴增。具有SEQ ID NO:12所示序列的反向引物用於HC(VH域)擴增,而具有SEQ ID NO:13所示序列的反向引物用於LC擴增。將PCR產物通過凝膠電泳進行分離,使用來自GE Healthcare Bio-Sciences的GFX PCR DNA&Gel Band Purification試劑盒進行提取,並使用Zero Blunt TOPO PCR Cloning試劑盒和化學感受態TOP10大腸桿菌(Life Technologies)克隆以供測序。使用來自Applied Biosystems的AmpliTaq Gold Master Mix和M13uni/M13rev引物對選擇的菌落進行菌落PCR。使用ExoSAP-IT酶混合物(USB)進行菌落PCR清掃(clean-up)。在MWG Biotech,Martinsried Germany處使用M13uni(-21)/M13rev(-29)測序引物進行測序。使用Vector NTI Advance 11程序(Life Technologies)對序列進行分析和注釋。所有試劑盒和試劑均按照製造商的說明書使用。
針對每個雜交瘤鑑定單個獨特的鼠κ型LC和單個獨特的鼠HC,亞類IgG1:mAb 2F3、mAb 2F22、mAb 2F45和mAb 1F91。mAb 1F56的LC和HC序列顯示,該抗體在序列上與mAb 1F91相同。可變重鏈和可變輕鏈序列(不包括前導肽序列)的胺基酸序列示於SEQ ID NO 4-11中。
抗體LC和HC表達載體的生成
生成了基於CMV啟動子的表達載體(pTT載體),以供mAb 2F22的小鼠-人嵌合形式的瞬時表達。pTT載體由Yves Durocher(Durocher等人.Nucleic Acid Research,2002)開發用於瞬時蛋白質表達。除了CMV啟動子外,基於pTT的載體還包含pMB1起點、EBV起點和Amp抗性基因。
生成了嵌合mAb 2F22輕鏈的表達載體,該嵌合mAb 2F22輕鏈攜帶鼠2F22輕鏈可變區和用於mAb 0294、Fab 0296和Fab 0295表達的人κ恆定區(SEQ ID NO:15)。生成了嵌合mAb 2F22重鏈的表達載體,該嵌合mAb 2F22重鏈攜帶鼠2F22重鏈可變區和用於mAb 0294表達的全長人IgG4(S241P)恆定區(SEQ ID NO:17)或用於Fab 0296表達(SEQ ID NO:14)或Fab 0295表達(SEQ ID NO:18)的截短的人IgG4恆定區。
生成了基於pTT的LC表達載體,以用於嵌合mAb 2F22抗體和抗體片段的瞬時表達。首先,使用對N-末端和C-末端序列具有特異性的引物,在2步反應中從原始TOPO測序克隆PCR擴增與mAb 2F22的VL域對應的區域。通過2步重疊PCR將原始鼠信號肽交換為人CD33信號肽。第一有義引物攜帶CD33信號肽序列的C-末端部分,並且第二有義引物含有用於克隆目的的HindIII限制位點、緊鄰ATG起始密碼子上遊的Kozak序列(5′-GCCGCCACC-3′)和CD33信號肽序列的N-末端部分。反義引物在VL/CL過渡序列中含有框內BsiWI限制位點。將擴增的片段克隆至含有人κCL域序列的線性化的基於pTT的載體中,隨後轉化至大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
生成了基於pTT的HC表達載體,以用於嵌合mAb 2F22和Fab 2F22片段的瞬時表達。嵌合mAb和Fab片段被分別稱為mAb 0294、Fab 0296和Fab 0295。首先,使用對N-末端和C-末端序列具有特異性的引物,在2步反應中從原始TOPO測序克隆PCR擴增與mAb 2F22的VH域對應的區域。通過2步重疊PCR將原始鼠信號肽交換為人CD33信號肽。第一有義引物攜帶CD33信號肽序列的C-末端部分,並且第二有義引物含有用於克隆目的的HindIII限制位點、緊鄰ATG起始密碼子上遊的Kozak序列(5′-GCCGCCACC-3′)和CD33信號肽序列的N-末端部分。反義引物在VH/CH過渡序列中含有框內NheI限制位點。為了生成全長HC表達載體(mAb 0294的HC),對產生的VH域PCR片段進行限制性消化,並將其克隆至含有人IgG4(S241P)恆定區序列的線性化的基於pTT的載體中。包含絲氨酸241向脯氨酸的IgG4鉸鏈突變,以通過消除半抗體的形成來穩定IgG4抗體。突變的鉸鏈位置在根據Kabat編號時被稱為S241P,或者在根據EU索引編號時被稱為S228P。
為了生成用於Fab 0296表達的截短的HC表達載體,對產生的VH域PCR片段進行限制性消化,並將其克隆至含有截短人IgG4恆定區序列的線性化的基於pTT的載體中。基於IgG4的HC如Fab 0296HC的序列中所見在鉸鏈區中在人IgG4鉸鏈賴氨酸殘基之後截短(SEQ ID NO:14)。隨後將克隆反應轉化到大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
為了生成用於Fab 0295表達的截短的HC表達載體,對產生的VH域PCR片段進行限制性消化,並將其克隆至含有第二截短人IgG4恆定區序列的線性化的基於pTT的載體中。用於Fab 0295的基於IgG4的HC如Fab 0295 HC的序列中所見在鉸鏈區中在人IgG4鉸鏈的第一半胱氨酸殘基之後截短(SEQ ID NO:18)。隨後將克隆反應轉化到大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
mAb和Fab片段的表達和純化
通過基於pTT的LC和HC表達載體的共轉染,在EXPI293F細胞(Life Technologies)的懸浮培養物中瞬時表達抗TFPI抗體和Fab片段。以下程序描述了用於適應懸浮的EXPI293F細胞的通用轉染方案。
EXPI293F轉染
1)首先製備DNA和轉染試劑的單獨稀釋液。
a)每ml細胞培養物使用總計1μg的載體DNA(0.5μg LC載體和0.5μg HC載體)。將DNA在Opti-MEM培養基(Gibco)中以50μl培養基/μg DNA稀釋,混合,並在室溫(23-25℃)下溫育5min。
b)以2.7μl/μg DNA的濃度使用ExpifectaminTM 293(Life Technologies)作為轉染試劑。將ExpifectaminTM溶液在Opti-MEM培養基(Gibco)中稀釋18.5倍,混合,並在室溫(23-25℃)下溫育5min。
2)混合DNA和ExpifectaminTM 293稀釋液,並使其在室溫(23-25℃)下溫育10min。
3)將DNA-ExpifectaminTM 293混合物直接添加至EXPI293F細胞培養物中。
a)在轉染時,EXPI293F培養物的細胞密度應為2.8-3.2×106個細胞/ml。
4)將轉染的細胞培養物轉移至在36.5℃、8%CO2和85-125rpm下的定軌搖床培養箱中。
5)轉染後18小時,添加5ul ExpifectaminTM 293Transfection Enhancer1/ml培養物和50μl ExpifectaminTM 293Transfection Enhancer2/ml培養物,並將培養物返回到在36.5℃、8%CO2和85-125rpm下的定軌搖床培養箱中。
6)轉染後5天,通過離心收穫細胞培養上清液,然後通過0.22μm PES過濾器單元(Coming)過濾。
通用純化方案
根據製造商的說明書,使用來自GE Healthcare的MabSelect SuRe樹脂,通過標準親和色譜法純化mAb變體。將純化的抗體用pH 7.2的PBS緩衝液進行緩衝液交換。
使用由GE Healthcare開發的KappaSelect樹脂,通過標準親和色譜法純化Fab片段。將純化的Fab片段用pH 7.2的PBS緩衝液進行緩衝液交換。通過SEC-HPLC進行質量評價和濃度確定。
實施例3:抗體分箱(binning)
使用生物層幹涉法(Fortebio Octet RED384儀器,PALL Life Sciences)測量TFPI抗體1F91、2F3、2F22、2F45、MBS532510(MyBioSource.com)、10R-T141A(Fitzgerald Industries International)與Bay 2A8-K95L之間對於與全長TFPIα結合的競爭。(Bay 2A8是於WO2010/017196中公開的2A8Fab的人IgG4(S241P)抗體變體。Bay 2A8-K95L是在HC CDR3中攜帶K95L置換(Kabat編號)的Bay 2A8的人IgG4(S241P)抗體變體(與WO2012/135671中公開的在「Fab B」中發現的相對於2A8的置換相同)。)使用1∶1.2mol mAb∶mol生物素-試劑(Thermo Scientific,目錄號21335)的比例或遵循製造商的說明書,用生物素在賴氨酸殘基上隨機標記mAb 1F91、mAb 2F3、mAb 2F22、mAb 2F45和Bay 2A8-K95L。在鏈黴親和素Fortebio sensortips(PALL Life Sciences)上捕獲生物素標記的抗體,然後結合人TFPIα,隨後結合mAb 1F91、mAb 2F3、mAb 2F22、mAb 2F45、Bay 2A8-K95L、MBS532510或10R-T141A。結果在表1中示出(黑色表示mAb競爭結合TFPI。灰色表示mAb不競爭結合TFPI)。
這些數據顯示抗體落入3個不同的箱元中,表明它們具有不同的結合表位。箱元1:mAb 1F91、MBS532510和10R-T141A。箱元2:mAb 2F3、2F22和2F45。箱元3:Bay2A8K95L。
表1:指定的TFPI抗體之間對於與TFPI結合的競爭
實施例4:與mAb 2F22的Fab片段複合的TFPI(1-79)的晶體結構
使用X射線晶體學,高解析度地確定由酸性N-末端區和Kunitz型蛋白酶抑制劑結構域1(KPI-1)組成的人TFPI(1-79)的N-末端部分(SEQ ID NO:16)與Fab片段——mAb 2F22的Fab 0296(SEQ ID NO:14和15)複合的3D結構。結果表明,該抗體能夠結合TFPI的KPI-1,和前面N-末端的一部分。所得的人TFPI表位殘基包含Leu 16、Pro 17、Leu 19、Lys 20、Leu 21、Met 22、Phe 25、Cys 35、Ala 37、Met 39、Arg 41、Tyr 56、Gly 57、Gly 58、Cys 59、Glu 60、Gly 61、Asn 62、Gln 63、Arg 65、Phe 66、Glu 67、Glu 71和Met 75(SEQ ID NO:2)。
材料與方法
為了結晶學目的,如實施例2所述生成了基於CMV啟動子的表達載體(pTT載體),用於瞬時表達與mAb 2F22抗體片段對應的Fab片段,以用於結晶學。
如實施例2所述,使mAb 2F22的Fab片段在EXPI293F細胞中以鼠-人嵌合形式Fab 0296(SEQ ID NO:14和15)表達,並通過使用KappaSelect樹脂的標準親和色譜法純化。
將包括人TFPI的N-末端部分和另外的N-末端連接的GSSGSSG標籤的人TFPI KPI-1(SEQ ID NO:16),以及由對應於SEQ ID NO:15的輕鏈和對應於SEQ ID NO:14的重鏈片段組成的Fab 0296(兩者均在磷酸鹽緩衝鹽水(PBS)緩衝液(2升水中4片,GIBCO,目錄號18912-014,Invitrogen Corporation)中),在TFPI物質稍微摩爾過量(1.1∶1)的情況下混合。然後利用分子量截止值為10,000Da的Amicon Ultra-4離心過濾器將複合物濃縮至約10.0mg/ml。採用來自TTP Lab Tech號:4150-05800的96孔TTP IQ板和每孔100μl沉澱劑溶液,通過坐滴技術(sitting drop-technique)使晶體生長。該沉澱劑溶液含有20%w/v PEG 3350、200mM甲酸鉀,並以3∶1的比例與蛋白質溶液混合。初始總液滴大小為200nl,數天後出現晶體。通過將1μl含有75%沉澱劑溶液和25%甘油的冷凍溶液混合物轉移至含有晶體的液滴中而製備用於低溫冷凍的晶體。使浸泡進行約2分鐘。然後將晶體捕獲,在液氮中快速冷凍,並在數據採集期間通過低溫N2氣流保持在100K的溫度下。在MAX-lab,Lund,Sweden處,在束線BL911-3下以解析度採集晶體學數據。通過XDS軟體包[Kabsch,W.,J.Appl.Crystallogr.,(1993),Vol.26,第795-800頁]進行數據的空間群確定、整合和縮放。空間群被確定為C2,並且同步加速器數據的晶胞參數被分別確定為89.01Q、66.660、106.β角為111.18°。解析度下的R-sym為8.4%,並且完整性為99.5%。在約的解析度下,獨特反射的強度/西格瑪(強度)的平均值等於2.0。
採用蛋白質資料庫(PDB)[Berman,H.M.等人,Nucleic Acids Res.,(2000),Vol.28,第235-242頁]中登錄編號為1NGZ[Yin,J.等人,Proc Natl Acad Sci U S A.2003年2月4日,(100),Vol.100第856-861頁]的Fab分子的坐標,使用分子置換(MR)方法進行結構確定。Fab分子被分成兩個結構域:可變域和恆定域,二者均在MR計算中用作搜索模型。使用CCP4軟體包CCP4[Collaborative Computational Project,N.,Acta crystallographica.Section D,Biological crystallography,(1994),Vol.50,第760-763頁]的Molrep軟體[Vagin,A.等人,J.Appl.Crystallogr.,(1997),Vol.30,第1022-1025頁]來尋找恆定和可變Fab域的位置。在MR步驟中未找到KPI-1結構域,但差異電子密度圖顯示了在該階段下KPI-1結構域分子的近似位置。通過CCP軟體包的DM軟體進行的電子密度改進,以及隨後使用ARP-wARP軟體[Langer,G.等人,Nat Protoc,(2008),Vol.3,第1171-1179頁][Murshudov,G.N.等人,Acta Crystallographica Section D Biological Crystallography,(2011),Vol.67,第355-367頁]進行的自動模型構建和相改進,給出了Fab 0296分子和KPI-1結構域結構以及在KPI-1結構域之前的N-末端部分的幾乎完整的結構。對於TFPI(SEQ ID NO:2),從15至77的殘基被包含在X射線模型中,該X射線模型除KPI-1結構域之外還包含KPI-1N-末端的一些殘基(殘基26-76)。對於Fab 0296片段,觀察到輕鏈殘基1至212和重鏈殘基1至221。進入使用Coot軟體程序[Emsley,P.等人,Acta Crystallogr.Sect.D-Biol.Crystallogr.,(2004),Vol.60,第2126-2132頁]對電子密度圖的計算機圖形檢查、模型校正和建立的過程,及隨後使用CCP4軟體包的軟體程序Refmac5[Murshudov,G.N.等人,Acta Crystallographica Section D Biological Crystallography,(2011),Vol.67,第355-367頁]的晶體學細化。循環該過程,直到無法對該模型進行進一步的顯著改進。在解析度下的所有數據的最終R-和R-free分別為0.192和0.220。
結果
通過CCP4程序套裝[Collaborative Computational Project,N.,Acta crystallographica.Section D,Biological crystallography,(1994),Vol.50,第760-763頁]的軟體程序Areaimol[Lee,B.等人,J Mol Biol,(1971),Vol.55,第379-400頁][Saff,E.B.等人,Math Intell,(1997),Vo1.19,第5-11頁]計算在成對相互作用中排除的平均面積給出了晶體結構的人TFPI片段/抗TFPI mAb 2F22Fab片段分子複合物。
使用Fab 0296與TFPI片段分子之間的截止距離,通過運行CCP4程序套裝[Bailey,S.,Acta Crystallogr.Sect.D-Biol.Crystallogr.,(1994),Vol.50,第760-763頁]的Contacts軟體來鑑定TFPI KPI-1(包括於晶體結構中觀察到的TFPI的N-末端部分)(SEQ ID NO:2)與Fab 0296(SEQ ID NO:14和15)的直接接觸。來自可溶性TFPI片段/Fab 0296複合物晶體結構的結果顯示在表2中。
發現所得的TFPI KPI-1(包括TFPI N-末端區、Fab 0296的表位)包含TFPI的以下殘基(採用SEQ ID NO:2的序列編號):Leu 16、Pro 17、Leu 19、Lys 20、Leu 21、Met 22、Phe 25、Cys 35、Ala 37、Met 39、Arg 41、Tyr 56、Gly 57、Gly 58、Cys 59、Glu 60、Gly 61、Asn 62、Gln 63、Arg 65、Phe 66、Glu 67、Glu 71和Met 75。從距離、電荷-電荷相互作用、氫鍵、極性和疏水相互作用以及低溶劑可及性評價,以下殘基似乎是表位的特別重要的殘基:Arg 41、Arg 65和Glu 67(SEQ ID NO.2)。
因此,mAb 2F22(由Fab 0296表示)的TFPI表位包含KPI-1結構域之前的殘基(包括短N-末端α-螺旋),KPI-1結構域的β-鏈1之前的環中的殘基,和β-鏈1起始端中的殘基。其還包含β-鏈2末端中的殘基,和β-鏈2與KPI-1的C-末端α-螺旋之間的殘基,以及KPI-1的C-末端α-螺旋內的殘基。
因此,結果顯示,Fab 0296與TFPI KPI-1及前面N-末端區的部分特異性結合,所以mAb 2F22也與它們特異性結合。
針對TFPI KPI-1的Fab 0296互補位包括重(H)鏈(SEQ ID NO:14,表2)的殘基Val 2、Phe 27、Tyr 32、Trp 52、Arg 53、Gly 54、Gly 55、Ser 56、Ile 57、Asp 58、Tyr 59、Ala 61、Met 64、Lys 97、Ser 99、His 100、Asn 102、Tyr 103、Val 104、Gly 105和Tyr 106,以及輕(L)鏈(SEQ ID NO:15,表2)的殘基Pro 31、Ala 32、Tyr 49、Ser 50、Asn 53、Tyr 55、Thr 56、Tyr 91、Thr 92、Ser 93和Tyr 94。
表2:來自TFPI(1-79)/Fab 0296複合物晶體結構的數據
結晶複合物的TFPI KPI-1,K鏈(SEQ ID NO:2)與Fab 0296的重鏈(H鏈)(SEQ ID NO:14)和Fab 0296的輕鏈(SEQ ID NO:15)的相互作用。採用的距離截止值。通過CCP4套件[Collaborative Computational Project,N.,Acta crystallographica.Section D,Biological crystallography,(1994),Vol.50,第760-763頁]的CONTACT計算機軟體程序鑑定接觸。在最後一列中,如通過CONTACT計算的,「***」表示在此接觸(距離<)處存在較大的氫鍵可能性,「*」表示較小的可能性(距離>)。空白表示該程序認為不存在氫鍵的可能性。氫鍵在供體與接受體之間是特異性的,通常較強,並且容易鑑定。
這些結果顯示,抗-TFPI 0296與TFPI KPI-1和前面N-末端的部分特異性結合。
實施例5:mAb 2021和mAb 2021變體的克隆和工程化
本實施例描述了抗-TFPI抗體mAb 2021及其變體的克隆和工程化。
使用來自Qiagen的RNeasy-Mini試劑盒從M-hTFPI 4F36A1B2雜交瘤細胞中提取總RNA,並將其用作cDNA合成的模板。使用來自Clontech的SMARTTM RACE cDNA擴增試劑盒在5′-RACE反應中合成cDNA。使用Phusion Hot Start聚合酶(Finnzymes)和作為正向引物包含在SMARTTM RACE試劑盒中的通用引物混合物(UPM),通過PCR進行隨後的HC和LC序列的靶擴增。對於HC(VH域)擴增,使用具有以下序列的反向引物:
5』-CCCTTGACCAGGCATCCCAG-3』(SEQ ID NO:12)
對於LC擴增,使用具有以下序列的反向引物:
5』-GCTCTAGACTAACACTCATTCCTGTTGAAGCTCTTG-3』(SEQ ID NO:13)
將PCR產物通過凝膠電泳進行分離,使用來自GE Healthcare Bio-Sciences的GFX PCR DNA&Gel Band Purification試劑盒進行提取,並使用Zero Blunt TOPO PCR Cloning試劑盒和化學感受態TOP10大腸桿菌(Invitrogen)克隆以供測序。使用來自Applied Biosystems的AmpliTaq Gold Master Mix和M13uni/M13rev引物在選擇的菌落上進行菌落PCR。使用ExoSAP-IT酶混合物(USB)進行菌落PCR清除(clean-up)。在Eurofins MWG Operon,Germany處使用M13uni(-21)/M13rev(-29)或T3/T7測序引物進行測序。使用VectorNTI程序對序列進行分析並注釋。所有試劑盒和試劑均按照製造商的說明書使用。
鑑定單個獨特的鼠κ型LC和單個獨特的鼠HC,亞類IgG1。
用於移植的抗-HzTFPI4F36A1B2的表達載體的生成
生成了一系列基於CMV啟動子的表達載體(pTT載體),用於在由Yves Durocher開發的基於HEK293-6E EBNA的表達系統(Durocher等人,Nucleic Acid Research,2002)中瞬時表達抗TFPI抗體/抗體片段。除了CMV啟動子外,基於pTT的載體還包含pMB1起點、EBV起點和Amp抗性基因。
基於克隆的鼠抗-TFPI4F36A1B2 VH和VL序列,通過在人種系序列上的CDR移植來設計抗TFPI4F36的人源化形式。根據上述抗體的人源化設計,合成(GENEART AG)了移植的HzTFPI4F36 VH和VL區的DNA序列。獲得具有基本最小CDR移植並且沒有其它回復突變的序列。構建體中包含相應的LC和HC種系前導肽序列,以及緊鄰ATG起始密碼子上遊的Kozak序列(5′-GCCGCCACC-3′)。
生成基於pTT的表達載體以用於將移植的HzTFPI4F36抗體瞬時表達為人κ/IgG4(S241P)同種型。在IgG4鉸鏈區中引入在位置241(根據Kabat編號,對應於根據EU編號系統的殘基228(Edelman G.M.等人,Proc.Natl.Acad.USA 63,78-85(1969))處的脯氨酸突變,以消除單體抗體片段的形成,即由一條LC和一條HC組成的「半抗體(half-antibodies)」的形成。
對於HC表達載體,通過對GENEART克隆載體的限制酶消化切下VH片段,並將其克隆至含有人IgG4(S241P)CH域序列的線性化的基於pTT的載體中,隨後轉化至大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。對於LC表達載體,通過對GENEART克隆載體的限制酶消化切下VL片段,並將其克隆至含有人κ CL域序列的線性化的基於pTT的載體中,隨後轉化至大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
用來分離mAb 2021的定點突變
在移植的HzTFPI4F36的輕鏈(LC)和重鏈(HC)中生成一系列人至小鼠反向突變(被稱為回復突變)。
進行定點突變以在移植的HzTFPI4F36 LC/HC構建體中的特定殘基處引入人至小鼠反向突變(以下稱為回復突變),以優化移植的構建體。通過兩種不同的方法引入突變:
1.使用來自Stratagene的定點或多位點定向突變試劑盒引入點突變和組合突變。試劑盒根據製造商的方案使用。
2.還使用標準的2步重疊PCR方法引入點突變以及產生組合突變。
將用於移植的HzTFPI4F36的LC和HC表達質粒用作突變的模板。通過DNA測序來驗證所有最終構建體的序列。
mAb 2021 HC的最終序列攜帶具有使得框架序列與原始小鼠FR2相同的4個回復突變的FR2區,和3個CDR2突變體,即,與原始的移植序列相比,總共7個HC回復突變(A40T、G42E、G44R、S49A、Y59F、A60P、K64Q)(根據Kabat編號)。LC序列為移植的HzTFPI4F36 LC序列。根據Kabat限定CDR和框架。
最終mAb 2021的VH和VL區的胺基酸序列分別列為SEQ ID NO:19和SEQ ID NO:20。
為改善表達產量,將HC和LC的原始信號肽序列(人種系序列)交換為人CD33信號肽(SP)。通過用含有Kozak元件(GCCGCCACC)、起始密碼子和CD33信號序列的引物(有義引物)以及含有終止密碼子和EcoRI限制位點的引物(反義引物)對HC或LC片段進行標準PCR擴增來交換信號肽序列。將擴增的片段克隆至線性化的基於pTT的載體中,並轉化至大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
mAb 2021的低親和力變體的生成
在抗TFPI4F36A1B2的人源化從而獲得mAb 2021的過程中,鑑定了許多低親和力變體。這些變體通過定點突變如上所述生成。mAb 2007代表了這樣的低親和力變體。與移植的HzTFPI4F36的序列相比,mAb 2007在VH域中攜帶單個A60P回復突變(根據Kabat編號)。該變體具有分別比mAb 2021低至少一個數量級的TFPI結合親和力。最初移植的HzTFPI4F36變體(mAb 2000)展現出比mAb 2021低約三個數量級的TFPI結合親和力。
如實施例6所述生成用於表達mAb 2021和mAb 2021變體的Fab片段的表達載體。
實施例6:雙特異性分子的Fab組分
對於Fab-Fab化學綴合物,生成了用於表達mAb 2021的截短的HC的表達載體。mAb 2021的克隆、人源化和表達於WO2010/072691中描述,其通過引用併入本文。如SEQ ID NO:23中所見,mAb 2021的基於IgG4的HC在鉸鏈區中在mAb 2021 HC的人IgG4恆定區中的第一半胱氨酸殘基之後截短。該截短留下可用於化學綴合的C-末端半胱氨酸。通過使用基於標準限制酶的克隆生成截短的序列,以將mAb 2021 HC的VH片段引入至含有截短的人IgG4 CH域序列的線性化的基於pTT的工具箱(toolbox)載體中。通過DNA測序來驗證所有最終構建體的序列。使用mAb 2021 LC載體和上述截短的HC載體(SEQ ID NO:24和SEQ ID NO:23)將mAb 2021的Fab片段表達為Fab 0088。用於Fab 0088的LC表達載體與如上所述生成的用於mAb 2021的LC表達相同,即通過限制酶消化從GENEART克隆載體上切下VL片段,並將其克隆至含有人κ CL域序列的線性化的基於pTT的載體中。還生成了用於表達mAb 2021的低親和力變體的重鍊表達載體。該mAb 2021變體源自mAb 2021的人源化,並且於WO2010/072691和實施例5中描述。為了組裝表達載體,採用標準的基於限制酶的克隆將與實施例5中描述的mAb 2007和mAb 2000的HC的VH域相對應的區域克隆至含有截短的人IgG4 CH域序列的線性化的基於pTT的工具箱載體中。基於IgG4的HC在IgG4鉸鏈中的第一個半胱氨酸之後截短,從而留下可用於化學綴合的C-末端半胱氨酸。通過DNA測序來驗證最終構建體的序列。mAb 2007和mAb 2000的低親和力Fab片段分別表示為Fab 0094(SEQ ID NO:21和SEQ ID NO:22)和Fab 0313(SEQ ID NO:25和SEQ ID NO:26)。Fab 0094和Fab 0313攜帶與Fab 0088相同的LC。
實施例7a:1,16-雙(2,5-吡咯二酮-1-基)-4,13-二氮雜-7,10-二氧雜-3,14-二氧代十六烷的合成
將3-(2,5-吡咯二酮-1-基)丙酸(3.0g,18mmol)溶於二氯甲烷(20ml)中。將溶液添加至預洗滌的固定的二環己基碳二亞胺樹脂(Novabiochem,產品號8.55029.0025,批次:S5092229 036,2.3mmol/g,10g)中,總體積為100ml。將混合物攪拌30分鐘。
在60分鐘內將1,8-二氨基-3,6-二氧雜辛烷(1.3g,8.9mmol)在二氯甲烷(50ml)中的溶液逐滴添加到攪拌的溶液中。將混合物攪拌2h。將溶液過濾並真空濃縮至幹。產量:3.4g淡棕色固體。
通過LCMS([M+H]+:451)和NMR(1H和13C)表徵產物。
將一部分製備的物質在乙酸酐中加熱至90攝氏度,持續3小時。將溶液冷卻,與水混合,冷凍並凍幹。
通過LCMS([M+Na]+:473)表徵產物。該物質用於以下兩個實施例中所述的綴合。
實施例7b:Fab 0088和Fab 0295與1,16-雙(2,5-吡咯二酮-1-基)-4,13-二氮雜-7,10-二氧雜-3,14-二氧代十六烷綴合以得到BiFab 9041
將Fab分子(每種Fab 4mg)的溶液在磷酸鹽緩衝鹽水(PBS)中調節至2.5mg/ml(50微摩爾)。將溶解在PBS中的雙(磺酸苯基)苯基膦二鉀二水合物鹽(CAS:151888-20-9,Sigma-Aldrich)溶液添加至Fab在PBS中的溶液中,直至所得濃度為50微摩爾Fab和200微摩爾的膦試劑。將溶液在環境溫度下溫育過夜。
使用Amicon Ultracel離心過濾器(MWCO 10kDa,Millipore A/S,Hellerup,Denmark)將Fab 0088用pH 5.0的15mM乙酸鈉緩衝液,1.0M NaCl進行緩衝液交換(終體積為1ml)。
使用Amicon Ultracel離心過濾器(MWCO 10kDa,Millipore A/S,Hellerup,Denmark)將Fab 0295用pH4.5的15mM乙酸鈉緩衝液進行緩衝液交換。終體積:300微升。
將1,16-雙(2,5-吡咯二酮-1-基)-4,13-二氮雜-7,10-二氧雜-3,14-二氧代十六烷(5mg)添加至還原的Fab蛋白質。將所得混合物在室溫下溫育50分鐘。
將樣品加載到HiTrap SP HP柱(1ml,在pH 4.5的15mM乙酸鈉緩衝液中預處理,GE Healthcare Europe GmbH,DK-2605,Denmark)上。將固定的蛋白質用pH 4.5的15mM乙酸鈉緩衝液(15個柱體積)洗滌,並用pH 5.0的15mM乙酸鈉緩衝液,1M NaCl洗脫。將兩種蛋白質溶液混合併使用Amicon Ultracel離心過濾器(MWCO 10kDa,MWCO 10kDa,Millipore A/S,Hellerup,Denmark)濃縮。終體積:100微升。
將混合物在環境溫度下溫育20小時。將溶液加載到已在pH7.3的10mM HEPES,150mM NaCl中預處理的Superdex200 16/60柱(GE Healthcare Europe GmbH,DK-2605,Denmark)上。使用所述緩衝液洗脫化合物BiFab 9041。將所選級分合併,濃縮,並使用SDS-PAGE分析、SEC-MALS、LCMS和Edman序列測定進行分析。
實施例7c:Fab 0313和Fab 0295與1,16-雙(2,5-吡咯二酮-1-基)-4,13-二氮雜-7,10-二氧雜-3,14-二氧代十六烷綴合以得到BiFAab 9042
將Fab分子(5mg Fab 0313和7mg Fab 0295)的溶液在磷酸鹽緩衝鹽水(PBS)中調節至2.5mg/ml(50微摩爾)。將溶解在PBS中的雙(磺酸苯基)苯基膦二鉀二水合物鹽(CAS號:151888-20-9,Sigma-Aldrich)溶液添加至Fab在PBS中的溶液中,直至所得濃度為50微摩爾Fab和200微摩爾的膦試劑。將溶液在環境溫度下溫育過夜。
使用Amicon Ultracel離心過濾器(MWCO 10 kDa,Millipore A/S,Hellerup,Denmark)將Fab 0313用pH 5.0的15mM乙酸鈉緩衝液,1.0M NaCl進行緩衝液交換(終體積為1ml)。
使用Amicon Ultracel離心過濾器(MWCO 10kDa,Millipore A/S,Hellerup,Denmark)將Fab 0295用pH 4.5的15mM乙酸鈉緩衝液進行緩衝液交換。終體積:300微升。
將1,16-雙(2,5-吡咯二酮-1-基)-4,13-二氮雜-7,10-二氧雜-3,14-二氧代十六烷(5mg)添加至還原的Fab蛋白質。將所得混合物在室溫下溫育50分鐘。
將樣品加載到HiTrap SP HP柱(1ml,在pH 4.5的15mM乙酸鈉緩衝液中預處理,GE Healthcare Europe GmbH,DK-2605,Denmark)上。將固定的蛋白質用pH 4.5的15mM乙酸鈉緩衝液(15個柱體積)洗滌,並用pH 5.0的15mM乙酸鈉緩衝液,1M NaCl洗脫。
將兩種蛋白質溶液混合,並使用Amicon Ultracel離心過濾器(MWCO 10kDa,MWCO 10kDa,Millipore A/S,Hellerup,Denmark)濃縮。終體積:100微升。
將混合物在環境溫度下溫育20小時。將溶液加載到已在pH 7.3的10mM HEPES,150mM NaCl中預處理的Superdex20016/60柱(GE Healthcare Europe GmbH,DK-2605,Denmark)上。使用所述緩衝液洗脫BiFab 9042化合物。將所選級分合併,濃縮,並使用SDS-PAGE分析、SEC-MALS和LCMS進行分析。
實施例8:結合相互作用分析
在通過表面等離子體共振實時測量分子相互作用的Biacore T200(GEHealthcare)上進行結合研究。實驗在25℃下進行,並且樣品在15℃下儲存在樣品室中。由Biacore報告的信號(RU,響應單位)與四個串聯流動池中各個傳感器晶片表面上的質量直接相關。根據製造商的說明書,將山羊抗Fabκ輕鏈抗體(Nordic Biosite)固定在CM4傳感器晶片的全部四個流動池上。通過將蛋白質稀釋至運行緩衝液(10mM Hepes 0.3M NaCl,5mM CaCl2,0.05%表面活性劑P20,10mg/ml BSA,pH 7.4)中來進行純化的雙特異性分子的捕獲,並將其注射在流動池2、3或4上,從而在流動池1中產生僅固定有抗Fab抗體的參考表面。雙特異性分子為雙特異性mAb 0421(SEQ ID NO:27、28和15、29)或BiFab 9041(SEQ ID NO:15、18、23、24)。通過在所有流動池上注射分析物(抗原)來進行TFPI FL(SEQ ID NO:1)、TFPI(1-79)和TFPI KPI-2蛋白的結合,以允許進行與不同經捕獲的雙特異性分子的結合相對於與參考表面的結合的比較分析。TFPI(1-79)片段跨越TFPI的N-末端和KPI-1結構域,具有N-末端連接的GSSGSSG標籤(SEQ ID NO:16)。TFPI KPI-2片段跨越TFPI的KPI-2結構域,具有C-末端HIS6標籤(SEQ ID NO:3)。將抗原連續稀釋至運行緩衝液中,以30μl/min注射240s,並使其解離600s或14000s。延長監測以接近1E-5s-1的kd緩慢解離的抗原的解離相對獲得解離速率的穩健值(robust value)是至關重要的。在每個分析物注射循環後,通過注射pH 2.1的10mM甘氨酸使CM4表面再生。該再生步驟將抗TFPI雙特異性分子和任何結合的抗原從固定的捕獲抗體表面上除去,並允許下一個相互作用樣品對的隨後結合。該再生程序沒有從晶片表面上除去直接固定的抗Fab捕獲抗體。
通過確定平衡解離常數(KD)來定量BiFab或雙特異性mAb與抗原之間的結合親和力,該平衡解離常數通過測量複合物形成和解離的動力學而確定。使用用於數據分析的Biacore評價軟體,通過將數據全局擬合至具有局部Rmax的1∶1Langmuir模型來檢索與單價複合物的結合和解離相對應的速率常數,如ka(結合速率)和kd(解離速率)。KD通過方程式KD=kd/ka與ka和kd相關。
通過雙參考(在數據分析之前在捕獲的雙特異性分子基礎上減去參考表面信號以及空白緩衝液注射)來處理結合曲線。這允許在樣品注射期間針對儀器噪聲、體積移位和漂移進行校正。
表3:TFPI蛋白與雙特異性分子結合的相互作用動力學
人TFPI(1-79)或KPI-2與不同雙特異性分子的結合常數ka(結合速率)、kd(解離速率)和KD(平衡解離常數)的測量結果。
在TFPI(1-79)的結合和TFPI KPI-2結合中發現BiFab 9041與mAb 0421沒有大的差異(表3)。與單個TFPI結構域相比,TFPI FL的結合顯示出不同的與兩種雙特異性分子的結合譜,因為結合似乎受到多價或親合力結合的影響(即使在低配體密度下),從而產生更高的表觀親和力。TFPI FL與mAb 0421和BiFab 9041的結合具有類似的結合曲線形狀,這表明通過經由兩個抗體片段的化學綴合而引入雙特異性結合(如同本實施例中使用的BiFab中一樣),實現了與雙特異性抗體相同的對結合譜的影響(參見圖3)。
實施例9:凝血酶生成試驗
在正常人血漿(CryoCheck Normal Plasma)中研究抗體對凝血酶生成的影響。通過再鈣化並添加1pM組織因子與4μM磷脂囊泡(PPP-Reagent low)而誘導凝血的啟動。通過添加100μg/ml綿羊抗人FVIII抗體(Haematologic Technologies Inc.,PAHFVIII-S)獲得A型血友病樣血漿。當指示時,向血漿中加入10-20nM外源重組人TFPIα(SEQ ID NO:1)。在來自Bachem(Bubendorf,Switzerland)的螢光底物Z-Gly-Gly-Arg-AMC.HCl(I-1140)轉化後,連續評價凝血酶活性。使用分別設定在368和460nm的激發和發射波長,在微量滴定板Fluorskan Ascent螢光計(Thermo Labsystems,Helsinki,Finland)中測量螢光。使用校準器來允許計算所形成的凝血酶的量並針對內濾效應和螢光底物消耗來校正所獲得的相對螢光單位。此外,減去凝血酶-α2-巨球蛋白複合物對底物轉化的貢獻。這些校正通過由Thrombinoscope BV(Maastricht,the Netherlands)提供的經校準的自動血栓圖(thrombogram)(CAT)計算機軟體自動進行。獲得產生凝血酶生成曲線的數據的一階導數,從而允許計算i)滯後時間,ii)曲線下總面積,內源性凝血酶電位(ETP),iii)凝血酶峰高度(峰值),iv)達到峰值的時間(ttPeak),和v)凝血酶生成的最大速率(速率)。
實施例10:TFPI(1-161)特異性抗TFPI抗體對TFPIα水平升高的人FVIII中和的血漿中組織因子誘導的凝血酶生成的影響
根據實施例9(圖1),在凝血酶生成試驗中研究了抗TFPI(1-79)抗體mAb 1F91、mAb 2F22、mAb 2F45和mAb 2F3以及TFPI KPI-2抗體mAb 2021對凝血酶生成的影響。來自圖1中的凝血酶生成的參數可參見表4。達到峰值的時間(ttPeak,min)和凝血酶峰(nM)都是n=3的平均值。將200nM抗體添加至補充有20nM TFPIα的FVIII中和的血漿中。
表4:凝血酶峰高(峰值)和達到峰值的時間(ttPeak)
自凝血酶生成曲線衍生的凝血酶峰高(峰值)和達到峰值的時間(ttPeak),該凝血酶生成曲線在補充有20nM TFPIα的FVIII中和的血漿中獲得。
一些但並非全部高親和力TFPI(1-79)抗體能夠部分恢復具有20nM TFPIα的A型血友病樣血漿中的凝血酶生成。表4中的數據提示,當循環中TFPI抗體的存在導致血漿TFPI濃度積累和增加時,需要更有效的TFPI抑制阻斷。
在根據實施例9的凝血酶生成試驗中研究了將每種抗TFPI(1-79)抗體與抗TFPI KPI-2mAb 2021組合對凝血酶生成的影響,並於圖2中示出。來自圖2中的凝血酶生成的參數可參見表5。達到峰值的時間(ttPeak,min)和凝血酶峰(nM)都是n=3的平均值。將100nM抗體添加至補充有20nM TFPIα和100nM mAb 2021的FVIII中和的血漿中。
表5:凝血酶峰高(峰值)和達到峰值的時間(ttPeak)
自凝血酶生成曲線衍生的凝血酶峰高(峰值)和達到峰值的時間(ttPeak),該凝血酶生成曲線在補充有20nM TFPIα的FVIII中和的血漿中獲得。將100nM的每種抗體添加至血漿中。
表5中的數據提示,通過中和參與TF/FVIIa/FXa/TFPI複合物穩定化的超過一個表位,獲得了更有效的TFPI抑制阻斷。
實施例11:抗TFPI BiFab對人A型血友病血漿中組織因子誘導的凝血酶生成的協同效應
根據實施例2和5-7製備了針對TFPI(1-79)、TFPI的KPI-2結構域的Fab片段和BiFab。將靶向TFPI(1-79)和KPI-2結構域的BiFab對凝血酶生成的影響與單獨或組合的相應未綴合Fab片段的影響進行比較。根據實施例9測量對FVIII中和的血漿和補充有10nM TFPIα的FVIII中和的血漿中的凝血酶生成的影響,並且數據於表6中列出。參數衍生自凝血酶生成曲線。達到峰值的時間(ttPeak,min)和凝血酶峰(nM)都是n=2的平均值。
表6:凝血酶峰高(峰值)和達到峰值的時間(ttPeak)
自凝血酶生成曲線衍生的凝血酶峰高(峰值)和達到峰值的時間(ttPeak),該凝血酶生成曲線在具有內源TFPI水平或補充有10nM TFPIα的FVIII中和的血漿中獲得。當使用FAb片段的組合時,每種片段被添加至100nM的濃度。
表6中的數據提示,通過中和TFPI上的超過一個表位,獲得了更有效的TFPI抑制阻斷。顯然,由於BiFab 9042與TFPI結合的親合力效應(實施例8)而獲得的高親和力導致了對凝血酶生成的協同效應,其高於通過將相應的兩個未綴合的Fab片段組合而獲得的效應。
實施例12:抗TFPI雙特異性抗體對人A型血友病血漿中組織因子誘導的凝血酶生成的影響
通過使用經由靶向TFPI(1-79)和TFPI的KPI-2區中的表位的抗體的Fc異二聚化而組裝的不對稱雙特異性抗體,可獲得對人A型血友病血漿中組織因子誘導的凝血酶生成的促凝血作用。
不對稱雙特異性抗體的生成
基於不對稱IgG形式,即通過Fc異二聚化,生成雙特異性TFPI(1-79)/KPI-2結合抗體。為了實現FC異二聚化,將一組相容的突變工程改造至mAb 2021(KPI-2結合)和mAb 2F22(TFPI(1-79)結合)的hIgG1變體的Fc區中。
通過限制酶消化從實施例5中描述的人源化mAb 2021的HC表達載體和HC表達載體mAb 2F22表達載體上切下VH片段,並將其克隆至含有人IgG1恆定區序列的線性化的基於pTT的工具箱載體中。隨後將克隆反應轉化到大腸桿菌中以供選擇。通過DNA測序來驗證最終構建體的序列。
為通過受控的Fab-臂交換來支持FC異二聚化,向mAb 2021的IgG1 HC的CH3域中引入單個賴氨酸至精氨酸的突變,其對應於SEQ ID NO:27的位置413。通過使用來自Stratagene的定點突變試劑盒的定點突變來引入該突變。通過DNA測序來驗證所有最終構建體的序列。mAb 2021的工程化IgG1形式如實施例2中所述在EXPI293F細胞中表達為mAb 0310(SEQ ID NO:27和28)。LC表達載體對應於如實施例5中所述的用於mAb 2021表達的載體。根據製造商的說明書,通過使用來自GE Healthcare的MabSelectSuRe樹脂的標準親和色譜法進行純化。將純化的抗體用pH 7.2的PBS緩衝液進行緩衝液交換。
向mAb 2F22的IgG1 HC的CH3域中引入匹配的苯丙氨酸至亮氨酸突變,其對應於SEQ ID NO:29中的位置409。通過使用來自Stratagene的定點突變試劑盒的定點突變來引入該突變。通過DNA測序來驗證所有最終構建體的序列。mA2 2F22的工程化IgG1形式如實施例2中所述在EXPI293F細胞中表達為mAb 0336(SEQ ID NO:15和29)。LC表達載體對應於如實施例2中所述的用於mAb 2F22表達的載體。根據製造商的說明書,通過使用來自GE Healthcare的MabSelectSuRe樹脂的標準親和色譜法進行純化。將純化的抗體用pH 7.2的PBS緩衝液進行緩衝液交換。
通過mAb2021(KPI-2結合)的工程化IgG1變體mAb 0310與mAb 2F22(TFPI(1-79)結合)的工程化IgG1變體mAb 0336之間的受控Fab-臂交換,製備了雙特異性TFPI(1-79)/KPI-2結合抗體。基本上根據公開的方法(Labrijn等人,PNAS,110,5145-5150(2013))製備該雙特異性抗體。簡言之,將mAb 0310(mAb 2021 IgG1 Lys至Arg變體SEQ ID NO:27)與mAb 0336(mAb 2F22 IgG1 Phe至Leu變體SEQ ID NO:29)以1∶1摩爾比(每種抗體1mg/ml)於PBS緩衝液中混合。通過在25mM 2-巰基乙胺(MEA)的存在下於37℃溫育該1∶1混合物90分鐘來選擇性地還原該抗體,以形成半抗體並允許Fab-臂交換。最後,在除去還原劑(滲濾至不含MEA的PBS緩衝液中)並在+5℃下儲存至少16小時後獲得自發重組裝和再氧化。觀察到由CH3域突變促進的異二聚化,效率超過90%。衍生自mAb 0310和0336的最終雙特異性抗體被命名為mAb 0421(SEQ ID NO:27、28和15、29)。
抗TFPI雙特異性抗體對人A型血友病血漿中組織因子誘導的凝血酶生成的影響
通過使用經由靶向TFPI(1-79)和TFPI的KPI-2區中的表位的抗體的Fc異二聚化而組裝的不對稱雙特異性抗體,可獲得對人A型血友病血漿中組織因子誘導的凝血酶生成的促凝血作用。根據實施例9於凝血酶生成試驗中研究了對FVIII中和的血漿和補充有10nM TFPIα的FVIII中和的血漿中凝血酶生成的影響,並且數據於表7中列出。參數衍生自凝血酶生成曲線。
表7:凝血酶峰高(峰值)和達到峰值的時間(ttPeak)
表7顯示,向正常血漿中添加抗人FVIII抗體以模擬A型血友病樣條件大大減少了凝血酶生成。凝血酶峰值(峰值)和達到峰值的時間(ttPeak)都受到影響。向FVIII中和的血漿中添加一些抗TFPI抗體,包括mAb 0310(mAb 2021的Fc工程化形式)、mAb 0336(抗TFPI mAb 2F22的Fc工程化形式)和mAb 0421(衍生自mAb 0310和mAb 0336的不對稱雙特異性抗體),有效地重新確立了凝血酶峰值(峰值)和達到峰值的時間(ttPeak)。據估計,獲得半數最大響應(EC50)所需的mAb 0421濃度為1.1nM(使用GraphPad PrismR軟體將表7中給出的2次實驗平均值的數據擬合至四參數劑量-響應曲線)。
此外,表7顯示,向FVIII中和的血漿中添加10nM全長TFPIα完全阻止了可測量的凝血酶生成,並且在這些條件下100nM針對TFPI(1-79)(mAb 0336)或KPI-2(mAb 0310)的高親和力抗體的添加不能完全消除TFPI抑制。相比之下,不對稱雙特異性抗體mAb 0421劑量依賴性地建立了穩定的凝血酶峰和縮短的達到峰值的時間(ttPeak),估計的EC50為9.5nM(單次實驗)。
表7中的數據提示,通過中和TFPI上的超過一個表位,獲得更有效的TFPI抑制阻斷。
實施例13:與TFPI KPI-2高親和力結合且與TFPI KPI-1低親和力結合的雙特異性分子
對於雙特異性TFPI(1-79)/KPI-2結合分子的某些應用,一個臂與TFPI KPI-2高親和力結合而另一個臂與TFPI(1-79)低親和力結合可能是有利的。
對應於mAb 2F22的Fab 0296(SEQ ID NO:14(重鏈)和15(輕鏈))以高親和力與TFPI(1-79)結合,並且可充當用於設計對TFPI(1-79)的親和力降低(與Fab 0296相比)的變體的模板。
如實施例4所述,Fab 0296的互補位已經確定,並且包括重鏈(SEQ ID NO:14)的殘基Val 2、Phe 27、Tyr 32、Trp 52、Arg 53、Gly 54、Gly 55、Ser 56、Ile 57、Asp 58、Tyr 59、Ala 61、Met 64、Lys 97、Ser 99、His 100、Asn 102、Tyr 103、Val 104、Gly 105和Tyr 106,以及輕鏈(SEQ ID NO:15)的殘基Pro 31、Ala 32、Tyr 49、Ser 50、Asn 53、Tyr 55、Thr 56、Tyr 91、Thr 92、Ser 93和Tyr 94。
Fab 0296的高親和力由在其上述互補位殘基與TFPI(1-79)上的表位殘基之間的幾種相互作用,例如靜電、極性、疏水和氫鍵相互作用的組合造成。因此,通過用不同的胺基酸殘基置換一個或多個互補位殘基,從而弱化或加強互補位-表位相互作用,來調節Fab 0296對TFPI(1-79)的結合親和力。
通過例如在單個位置的保守取代、在數個位置的保守取代或在一個或多個位置的保守性較低的取代而不同程度地削弱Fab 0296對TFPI(1-79)的親和力。
除了在重鏈C-末端處的額外半胱氨酸外,Fab 0295(SEQ ID NO:18(重鏈)和15(輕鏈))與Fab 0296相同,並且其具有與Fab 0296相同的互補位。因此,通過使用其中單個或多個互補位殘基已被取代的Fab 0295的變體,並如實施例7所述將其與KPI-2結合Fab綴合,來製備對TFPI(1-79)具有低親和力的雙特異性TFPI(1-79)/KPI-2結合分子。KPI-2結合Fab可為例如Fab 0088,但也可為其它KPI-2結合Fab片段。
通過使用與本實施例中描述的Fab片段對應的全長抗體,根據實施例12中描述的方法獲得相應的雙特異性抗體是可能的。
雖然本文已經示出和描述了本發明的某些特徵,但是本領域普通技術人員將會想到許多修改、替換、改變和等同物。因此,應當理解,所附權利要求旨在覆蓋落入本發明真正精神內的所有這樣的修改和改變。
序列表
諾和諾德股份有限公司
能夠結合組織因子途徑抑制物(1-161)上的兩個表位的抗體
140072
29
PatentIn version 3.5
1
276
PRT
智人
1
Asp Ser Glu Glu Asp Glu Glu His Thr Ile Ile Thr Asp Thr Glu Leu
1 5 10 15
Pro Pro Leu Lys Leu Met His Ser Phe Cys Ala Phe Lys Ala Asp Asp
20 25 30
Gly Pro Cys Lys Ala Ile Met Lys Arg Phe Phe Phe Asn Ile Phe Thr
35 40 45
Arg Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Gln Asn
50 55 60
Arg Phe Glu Ser Leu Glu Glu Cys Lys Lys Met Cys Thr Arg Asp Asn
65 70 75 80
Ala Asn Arg Ile Ile Lys Thr Thr Leu Gln Gln Glu Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Thr Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Met Asn Asn Phe Glu Thr Leu Glu Glu Cys Lys
130 135 140
Asn Ile Cys Glu Asp Gly Pro Asn Gly Phe Gln Val Asp Asn Tyr Gly
145 150 155 160
Thr Gln Leu Asn Ala Val Asn Asn Ser Leu Thr Pro Gln Ser Thr Lys
165 170 175
Val Pro Ser Leu Phe Glu Phe His Gly Pro Ser Trp Cys Leu Thr Pro
180 185 190
Ala Asp Arg Gly Leu Cys Arg Ala Asn Glu Asn Arg Phe Tyr Tyr Asn
195 200 205
Ser Val Ile Gly Lys Cys Arg Pro Phe Lys Tyr Ser Gly Cys Gly Gly
210 215 220
Asn Glu Asn Asn Phe Thr Ser Lys Gln Glu Cys Leu Arg Ala Cys Lys
225 230 235 240
Lys Gly Phe Ile Gln Arg Ile Ser Lys Gly Gly Leu Ile Lys Thr Lys
245 250 255
Arg Lys Arg Lys Lys Gln Arg Val Lys Ile Ala Tyr Glu Glu Ile Phe
260 265 270
Val Lys Asn Met
275
2
161
PRT
人工序列
TFPI (1-161)
2
Asp Ser Glu Glu Asp Glu Glu His Thr Ile Ile Thr Asp Thr Glu Leu
1 5 10 15
Pro Pro Leu Lys Leu Met His Ser Phe Cys Ala Phe Lys Ala Asp Asp
20 25 30
Gly Pro Cys Lys Ala Ile Met Lys Arg Phe Phe Phe Asn Ile Phe Thr
35 40 45
Arg Gln Cys Glu Glu Phe Ile Tyr Gly Gly Cys Glu Gly Asn Gln Asn
50 55 60
Arg Phe Glu Ser Leu Glu Glu Cys Lys Lys Met Cys Thr Arg Asp Asn
65 70 75 80
Ala Asn Arg Ile Ile Lys Thr Thr Leu Gln Gln Glu Lys Pro Asp Phe
85 90 95
Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys Arg Gly Tyr Ile Thr Arg
100 105 110
Tyr Phe Tyr Asn Asn Gln Thr Lys Gln Cys Glu Arg Phe Lys Tyr Gly
115 120 125
Gly Cys Leu Gly Asn Met Asn Asn Phe Glu Thr Leu Glu Glu Cys Lys
130 135 140
Asn Ile Cys Glu Asp Gly Pro Asn Gly Phe Gln Val Asp Asn Tyr Gly
145 150 155 160
Thr
3
66
PRT
人工序列
TFPI KPI-2構建體
3
Gln Glu Lys Pro Asp Phe Cys Phe Leu Glu Glu Asp Pro Gly Ile Cys
1 5 10 15
Arg Gly Tyr Ile Thr Arg Tyr Phe Tyr Asn Asn Gln Thr Lys Gln Cys
20 25 30
Glu Arg Phe Lys Tyr Gly Gly Cys Leu Gly Asn Met Asn Asn Phe Glu
35 40 45
Thr Leu Glu Glu Cys Lys Asn Ile Cys Glu Asp Gly His His His His
50 55 60
His His
65
4
121
PRT
小鼠
4
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ser Ser Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Thr Asp Phe Asn Ala Ala Phe Met
50 55 60
Ser Arg Val Ser Phe Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
5
107
PRT
小鼠
5
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Thr Val Gly
1 5 10 15
Asp Lys Val Ser Ile Pro Cys Lys Ala Ser Glu Asn Val Gly Ala Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Thr Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Ile Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Arg Thr Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Thr Asn Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
6
121
PRT
小鼠
6
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
7
107
PRT
小鼠
7
Gly Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Thr Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Ser Val Gly Pro Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Phe Thr Leu Thr Ile Ser Asn Leu His Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Thr Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
8
121
PRT
小鼠
8
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Ser Cys Thr Val Ser Gly Phe Ser Leu Thr Gly Tyr
20 25 30
Gly Val His Trp Ile Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser
115 120
9
107
PRT
小鼠
9
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Thr Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Gly Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Glu Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Arg Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Met Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Thr Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105
10
115
PRT
小鼠
10
Asp Val Gln Leu Gln Glu Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1 5 10 15
Ser Leu Ser Leu Thr Cys Thr Val Ser Gly Tyr Ser Ile Thr Ser Asp
20 25 30
Tyr Ala Trp Asn Trp Ile Arg Gln Phe Pro Gly Asn Lys Leu Glu Trp
35 40 45
Met Gly Tyr Ile Ser Tyr Ser Gly Ser Thr Ser Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Ile Ser Ile Thr Arg Asp Thr Ser Lys Asn Gln Phe Phe
65 70 75 80
Leu Gln Leu Asn Ser Val Thr Thr Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Ala Arg Trp Ala Tyr Asp Gly Pro Trp Gly Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ala
115
11
107
PRT
小鼠
11
Gln Ile Val Leu Ser Gln Ser Pro Ala Leu Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser His Met
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Thr Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Phe Thr
85 90 95
Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Arg
100 105
12
20
DNA
人工序列
引物
12
cccttgacca ggcatcccag 20
13
36
DNA
人工序列
引物
13
gctctagact aacactcatt cctgttgaag ctcttg 36
14
222
PRT
人工序列
Fab 0296的截短的鼠-人嵌合重鏈
14
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
210 215 220
15
213
PRT
人工序列
mAb 0294、Fab 0296和Fab 0295和mAb 0336的鼠-人嵌合輕鏈
15
Gly Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Thr Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Ser Val Gly Pro Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Ala Asp Phe Thr Leu Thr Ile Ser Asn Leu His Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Thr Ser Tyr Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
16
86
PRT
人工序列
標記的TFPI KPI-1/N-末端(TFPI (1-79))
16
Gly Ser Ser Gly Ser Ser Gly Asp Ser Glu Glu Asp Glu Glu His Thr
1 5 10 15
Ile Ile Thr Asp Thr Glu Leu Pro Pro Leu Lys Leu Met His Ser Phe
20 25 30
Cys Ala Phe Lys Ala Asp Asp Gly Pro Cys Lys Ala Ile Met Lys Arg
35 40 45
Phe Phe Phe Asn Ile Phe Thr Arg Gln Cys Glu Glu Phe Ile Tyr Gly
50 55 60
Gly Cys Glu Gly Asn Gln Asn Arg Phe Glu Ser Leu Glu Glu Cys Lys
65 70 75 80
Lys Met Cys Thr Arg Asp
85
17
448
PRT
人工序列
單克隆抗體mAb 0294的鼠-人嵌合重鏈
17
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
260 265 270
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Lys
435 440 445
18
227
PRT
人工序列
Fab 0295的截短的鼠-人嵌合重鏈
18
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys
225
19
121
PRT
人工序列
mAb 2021的重鏈可變域(VH)
19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser
115 120
20
113
PRT
人工序列
mAb 2021的輕鏈可變域(VL)
20
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg
21
227
PRT
人工序列
Fab 0094的重鏈(HC)
21
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys
225
22
219
PRT
人工序列
Fab 0094的輕鏈(LC)
22
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
23
227
PRT
人工序列
Fab 0088的重鏈(HC)
23
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys
225
24
219
PRT
人工序列
Fab 0088的輕鏈(LC)
24
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
25
227
PRT
人工序列
Fab 0313的重鏈(HC)
25
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val Asp His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys Tyr Gly
210 215 220
Pro Pro Cys
225
26
219
PRT
人工序列
Fab 0313的輕鏈(LC)
26
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
27
451
PRT
人工序列
mAb 0310的重鏈(HC)
27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asn Tyr
20 25 30
Ala Met Ser Trp Val Arg Gln Thr Pro Glu Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Ser Gly Ser Tyr Ser Tyr Phe Pro Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Leu Gly Gly Tyr Asp Glu Gly Asp Ala Met Asp Ser Trp Gly
100 105 110
Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
28
219
PRT
人工序列
mAb 0310的輕鏈(LC)
28
Asp Ile Val Met Thr Gln Thr Pro Leu Ser Leu Ser Val Thr Pro Gly
1 5 10 15
Gln Pro Ala Ser Ile Ser Cys Lys Ser Ser Gln Ser Leu Leu Glu Ser
20 25 30
Asp Gly Lys Thr Tyr Leu Asn Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Val Ser Ile Leu Asp Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Leu Gln Ala
85 90 95
Thr His Phe Pro Gln Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
29
451
PRT
人工序列
mAb 0336的重鏈(HC)
29
Gln Val Gln Leu Lys Gln Ser Gly Pro Gly Leu Val Gln Pro Ser Gln
1 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Thr Asn Tyr
20 25 30
Gly Val His Trp Val Arg Gln Ser Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Ala Val Ile Trp Arg Gly Gly Ser Ile Asp Tyr Asn Ala Ala Phe Met
50 55 60
Ser Arg Leu Ser Ile Thr Lys Asp Asn Ser Lys Ser Gln Val Phe Phe
65 70 75 80
Lys Met Asn Ser Leu Gln Ala Asp Asp Thr Ala Ile Tyr Tyr Cys Ala
85 90 95
Lys Asn Ser His Gly Asn Tyr Val Gly Tyr Ala Met Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Leu Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450