CRM197和相關蛋白的表達和純化的製作方法
2023-04-23 14:56:22 5
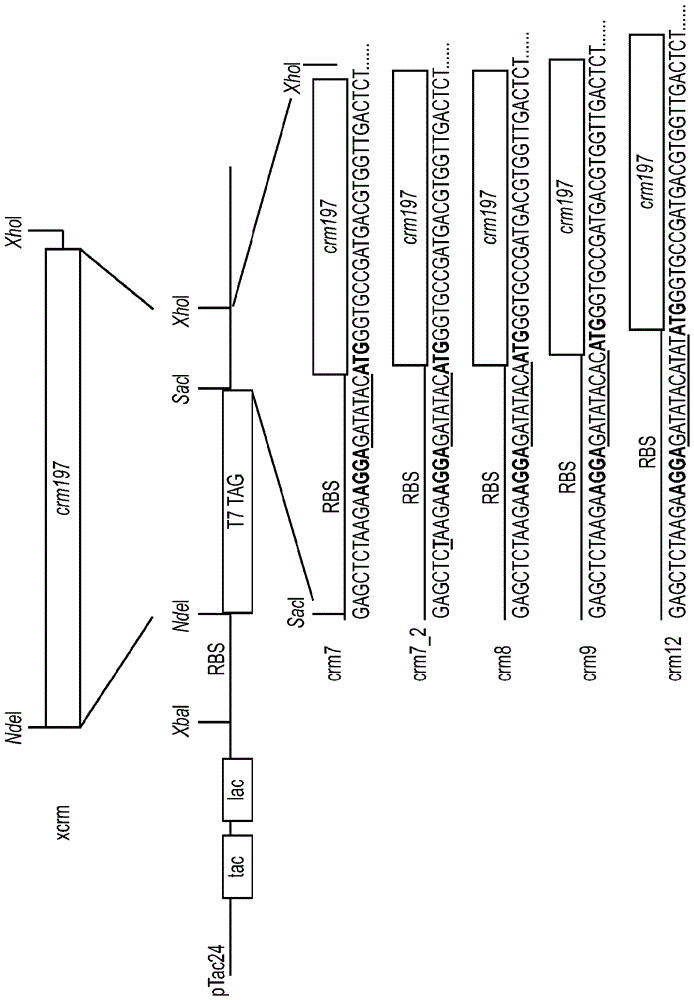
本申請要求2014年1月31日提交的相同標題的美國臨時申請號61/934,377的優先權,其整體通過引用特別結合到本文中。發明背景1.發明領域本發明涉及在細菌宿主中生產重組蛋白的領域。具體而言,本發明涉及用於從大腸桿菌獲得高水平的可溶性重組CRM197蛋白的生產方法。本發明還涉及CRM197的純化和表徵方法以及通過所述方法生產的CRM197的用途。2.背景描述白喉毒素(DT)是一種由白喉棒狀桿菌(Corynebacteriumdiphtheriae)的致病菌株合成和分泌的蛋白質性外毒素。這些致病菌株包含攜帶毒素基因的噬菌體溶素原。白喉毒素是一種ADP-核糖基化的酶,其作為535個殘基的酶原分泌和由胰蛋白酶-樣蛋白酶加工,並釋放兩個片段(A和B)。片段A使用NAD作為底物,催化在煙醯胺環和N-核糖之間的N-糖苷鍵的裂解和介導ADP-核糖共價轉移(ADPRT活性)至延伸因子EF-2的修飾的組氨酸715(白喉醯胺)。該翻譯後白喉醯胺修飾使EF-2失活,停止蛋白合成和導致細胞死亡。DT的A片段(亦稱為C結構域)帶有催化活性位點,並且是中毒的最後一步所需的唯一毒素片段。B片段上攜帶的R結構域介導與宿主細胞表面上的受體的結合,並且也在B片段上攜帶的T結構域促進片段A的pH-依賴性轉移至細胞質。富含精氨酸的二硫鍵連接的環連接片段A與片段B(或者結構域C與結構域TR)。在該鏈在位置186上蛋白水解裂解後,該鏈間二硫鍵是在這兩個片段之間唯一的共價連接。各種無毒性和部分毒性的免疫學交叉反應形式的白喉毒素(CRM或交叉反應材料)的分離導致發現CRM197(Uchida等JournalofBiologicalChemistry248,3845-3850,1973;亦參見Giannini等NucleicAcidsRes.1984年5月25日;12(10):4063-9)。優選地,CRM可具有包含整個DT或其一部分的任何大小和組成。CRM197是一種主要酶失活和無毒性形式的白喉毒素,其包含單個胺基酸置換G52E。該突變導致NAD-結合位點之前的活性位點環的內在柔韌性,和降低CRM197結合NAD的能力並消除DT的有毒性質(Malito等ProcNatlAcadSciUSA109(14):5229-342012)。如同DT一樣,CRM197具有兩個二硫鍵。一個二硫鍵連接Cys186與Cys201,其連接片段A與片段B。第二個二硫鍵在片段B內連接Cys461與Cys471。DT和CRM197兩者均具有片段A-相關的核酸酶活性(Bruce等Proc.Natl.Acad.Sci.USA87、2995-8、1990)。許多抗原具有差的免疫原性,尤其是在嬰兒中,除非與蛋白化學連接(「綴合」),從而形成綴合物或綴合物疫苗。這些綴合物疫苗的蛋白組分也稱為「載體蛋白」。CRM197通常用作蛋白-糖和半抗原-蛋白綴合物的載體蛋白。作為載體蛋白,CRM197相對於白喉類毒素以及其它類毒素蛋白具有許多優點,其中許多已被證明(ShinefieldVaccine,28:4335,2010;Broker等Biologicals,39:1952011)。例如,因為CRM197是遺傳上去毒性的,它保留較大的賴氨酸(其用於綴合但被化學類毒素化封閉)補體。已經證明CRM197是肺炎鏈球菌莢膜多糖的有效的載體蛋白,如通過PREVNAR™(Pfizer)(一種由與CRM197化學連接的至多13種莢膜多糖組成的疫苗)的成功所證明的。還有證據表明,與破傷風類毒素相比,存在較少的載體誘導的免疫反應抑制,尤其是當存在許多單獨多糖與相同的載體蛋白連接時。CRM197和天然DT對白喉毒素受體(DTR)具有類似的親和力,所述受體與HB-EGF前體pro-HB-EGF具有相同的胺基酸序列(Mitamura等J.Biol.Chem.272(43):27084-90、1997)。CRM197結合可溶形式的HB-EGF,以及膜形式pro-HB-EGF,和通過阻止其與EGF受體結合而抑制HB-EGF有絲分裂作用。因此CRM197也可能在癌症療法中具有有前景的作用(Miyamoto等AnticancerRes.Nov-Dec27(6A):3713-21,2007)。CRM197已經在原始宿主棒狀桿菌中生產,但收率低,通常<50mg/L,並且另外地,與例如大腸桿菌相比,棒狀桿菌生長相對緩慢。存在專有的棒狀桿菌菌株,其已被工程改造以較高水平生產CRM197(美國專利號5,614,382)。CRM197還已經在專有的螢光假單胞菌(Psuedomonasfluorescens)菌株中表達和以高水平表達。在大腸桿菌中生產CRM197將是有利的,因為大腸桿菌是BL1水平的生物,其在培養和繁殖方面廉價。在大腸桿菌中生產CRM197主要產生不溶性包涵體(通常不溶),其然後需要困難的再摺疊過程,導致收率低(EP20100742260)或具有額外的肽序列(標籤)(JBiotechnol.2010Dec20;156(4):245-52,OverexpressionandpurificationoftherecombinantdiphtheriatoxinvariantCRM197inEscherichiacoli.StefanA,ContiM,RubboliD,RavagliL,PrestaE,HochkoepplerA)。尚未報導適合於大量蛋白生產的用於在大腸桿菌中過量表達可溶性無標籤CRM197的方法。因此,對以高效和成本有效方式生產CRM197的更好方法存在需要。發明概述本發明克服了與當前策略和設計有關的問題和缺點,提供了用於生產CRM的新的組合物和方法。本發明的一個實施方案涉及生產整個CRM蛋白或其一部分的方法,包括:提供包含表達載體的重組細胞,所述表達載體包含與多順反子基因序列功能連接的誘導型啟動子,其中至少一個順反子編碼CRM蛋白;誘導表達載體生產CRM蛋白;和分離表達的CRM蛋白。優選地,重組細胞的一種或多種二硫化物還原酶的活性降低,和還優選地,各個順反子包含核糖體結合位點和起始密碼子。優選地,多順反子基因序列在一個或多個核糖體結合位點和一個或多個起始密碼子之間包含至少一個間隔基。優選地,由細胞表達的CRM蛋白是可溶性的,和還優選地,表達的CRM蛋白是細胞內的、周質的或分泌的。優選地,重組細胞在約15°C至約32°C的溫度下增殖,和還優選地,CRM蛋白通過色譜法從細胞分離。優選的色譜介質包括例如硫酸葡聚糖樹脂、凝膠樹脂、活性硫酸化樹脂、磷酸鹽樹脂、肝素樹脂或肝素-樣樹脂。本發明的另一實施方案包括通過本發明的方法分離的CRM蛋白。本發明的另一實施方案涉及生產整個CRM蛋白或其一部分(例如優選地CRM197)的方法,包括:提供包含表達載體的重組細胞,其中與未修飾的重組細胞相比,重組細胞已被修飾以改變細胞質的氧化還原狀態至更加氧化的狀態,和表達載體包含與CRM編碼序列化學連接的誘導型啟動子、在核糖體結合位點和ATG密碼子之間的間隔基序列、CRM編碼序列上遊的表達增強子區;誘導表達載體生產CRM蛋白;和分離表達的CRM蛋白。重組細胞可以是真核細胞或原核細胞。優選地,重組細胞是原核細胞,例如大腸桿菌細胞或大腸桿菌的衍生菌或菌株。優選地,重組細胞修飾包括一種或多種二硫化物還原酶的活性降低,所述二硫化物還原酶例如氧化還原酶、二氫葉酸還原酶、硫氧還蛋白和硫氧還蛋白還原酶、蛋白還原酶或穀胱甘肽還原酶的一種或多種。優選地,與非-重組細胞相比,一種或多種二硫化物還原酶的活性降低改變重組細胞的細胞質的氧化還原狀態至氧化狀態。優選地,CRM編碼序列編碼一個或多個CRM表位、CRM肽序列、CRM結構域或其組合。優選地,間隔基包含多於9個核苷酸或少於9個核苷酸,例如5-20個核苷酸。優選地,表達增強子包含CRM編碼序列上遊的核糖體結合位點和ATG密碼子。優選地,由細胞表達的CRM蛋白是可溶性的,和是細胞內的、周質的或分泌的。優選地,重組細胞在約15°C至約32°C的溫度下增殖。優選地,CRM蛋白通過色譜法從細胞分離,所述色譜法包括硫酸葡聚糖樹脂、活性硫酸鹽樹脂、磷酸鹽樹脂、肝素樹脂或肝素-樣樹脂作為優選的色譜介質。本發明的另一實施方案涉及通過本發明的方法分離的CRM蛋白。優選地,分離的CRM蛋白是綴合的,和綴合的CRM蛋白作為疫苗配製。本發明的另一實施方案涉及生產整個CRM蛋白或其一部分(例如從CRM編碼序列生產的蛋白或肽,所述編碼序列編碼一個或多個CRM表位、CRM肽序列、CRM結構域或其組合,和優選地CRM197)的方法,包括提供包含表達載體的重組細胞,其中所述表達載體包含與EES編碼序列(其前面有核糖體結合位點)功能連接的啟動子;從CRM編碼序列(其前面有核糖體結合位點)表達CRM蛋白;和分離表達的CRM蛋白。優選地,重組細胞是原核或真核細胞,和優選地,原核細胞是大腸桿菌細胞或大腸桿菌的衍生菌或菌株。優選地,啟動子是組成型或誘導型的。優選地,與未修飾的重組細胞相比,重組細胞被修飾以改變細胞質的氧化還原狀態至更加氧化的狀態。優選地,修飾的重組細胞的一種或多種硫醇-二硫化物氧化還原酶和/或參與硫氧還蛋白和谷氧還蛋白系統的酶(例如,硫氧還蛋白、硫氧還蛋白還原酶、穀胱甘肽還原酶)的活性降低。優選地,表達載體包含受核糖體結合位點控制的表達增強子序列(例如SEQID15)或例如包括CRM編碼序列上遊的核糖體結合位點上遊的T7標籤序列。本發明的另一實施方案涉及分離和/或純化CRM蛋白的方法,包括用加樣緩衝液加載CRM蛋白至包含樹脂的色譜柱上,其中樹脂優選地是硫酸葡聚糖樹脂、活性硫酸鹽樹脂、磷酸鹽樹脂、肝素樹脂或肝素-樣樹脂;用一種或多種洗滌緩衝液洗滌樹脂;和用洗脫緩衝液從樹脂洗脫CRM蛋白。優選地,加樣緩衝液和洗滌緩衝液是或包含相同的組分,並且其量相同或類似。優選地,加樣緩衝液和一種或多種洗滌緩衝液是低電導率緩衝液,例如Tris-HCl、HEPES、磷酸鈉緩衝液,電導率為約10mS/cm或更小(例如1mS/cm、2mS/cm、3mS/cm、4mS/cm、5mS/cm、6mS/cm、7mS/cm、8mS/cm、9mS/cm)。優選地,洗脫緩衝液是高電導率緩衝液,例如含有添加的鹽例如NaCl或KCl的緩衝液,電導率為約10mS/cm或更大(例如12mS/cm、14mS/cm、15mS/cm、20mS/cm、25mS/cm、30mS/cm、40mS/cm、50mS/cm、60mS/cm、70mS/cm、80mS/cm、90mS/cm、100mS/cm或更大)。本發明的另一實施方案涉及表徵白喉毒素或CRM蛋白的摺疊的方法,包括:使白喉毒素或CRM蛋白接觸HB-EGF;測定白喉毒素或CRM蛋白與HB-EGF結合的量;和通過測定的結合的量,測定白喉毒素或CRM蛋白的摺疊,其中結合表示正確摺疊。優選地,白喉毒素或CRM包含受體結合結構域。優選地,CRM蛋白包含CRM197。還優選地,白喉毒素或CRM蛋白和/或HB-EGF的至少一種結合到固體載體。優選地,白喉毒素或CRM蛋白與HB-EGF結合的量通過ELISA測定,和結合HB-EGF的CRM蛋白在PBS中是可溶性的。本發明的另一實施方案包含表達載體,其包含啟動子和兩個或更多個順反子(至少一個編碼蛋白),其中至少一個順反子編碼CRM蛋白和各個順反子具有核糖體結合位點和起始密碼子。優選地,表達載體在核糖體結合位點和起始密碼子之間進一步包含間隔基,和還優選地,間隔基包含5-20個核苷酸。優選地,間隔基不包含9個核苷酸。本發明的其它實施方案和優點在之後的詳述中部分地描述,和部分地,可從該詳述中顯而易見,或者可從本發明的實施中獲知。附圖簡述圖1:載體構建體crm7(SEQIDNO10)、crm7_2(SEQIDNO11)、crm8(SEQIDNO12)、crm9(SEQIDNO13)和crm12(SEQIDNO14)的示意圖,其中核糖體結合位點(RBS)和起始密碼子(ATG)兩者均用粗體表示,間隔基序列用下劃線表示。圖2:在用2µg/ml的pn6B包被的immunotech板上小鼠抗-pn6B血清的滴定結果。圖3:在pn14包被的板上抗-pn14血清的滴定結果。發明詳述可溶性的完整重組CRM最初在蛋白酶-缺陷的大腸桿菌中生產(Bishai等1987)。然而,蛋白生產量非常低。隨後,CRM197在大腸桿菌細胞中作為包涵體(StefanA等JBiotechnol.Dec20;156(4):245-52,2010;國際申請公開號WO2011/126811,中國專利申請號200610042194)或作為通過信號肽引導到周質的可溶性蛋白(國際申請公開號WO2011/042516)生產。大腸桿菌的周質是一種氧化環境,其允許形成二硫鍵。CRM197具有兩個二硫鍵,其對於正確摺疊和功能以及對於蛋白可溶性而言可能是重要的。已經令人驚訝地發現,可溶性重組CRM蛋白的單一未裂解鏈可從微生物在細胞內和以商業化的量快速生產,其後大量分離和/或純化,並保持可溶。CRM在磷酸鹽緩衝鹽水(PBS,pH7.5)和其它類似的緩衝液中是可溶性的,並且在該緩衝液和其它緩衝液中可濃縮至大於5mg/ml,同時保持可溶。儘管在Cornybacter中表達的CRM可在這些緩衝液中濃縮,但在假單胞菌中產生和在周質中表達的CRM不能容易地在這些相同的緩衝液中濃縮(PfenexInc.,SanDiego,CA)。與周質表達相比,細胞內表達的其它優點是實現更高表達水平,這是因為與細胞內空間相比,周質空間有限。優選生產的CRM蛋白是全長或部分區域,例如肽、單一或多個結構域或表位和從天然CRM編碼序列表達的任何特定區域,其包括已被一個或多個缺失、取代和/或添加(例如保守或非保守的)修飾的CRM序列,和已被促進在宿主生物中的表達的另外序列(例如一個或多個啟動子、起始密碼子和翻譯因子、核糖體或聚合酶結合位點)修飾的CRM序列。優選的CRM蛋白是CRM197。可溶性的和不作為細胞的不溶性包涵體另外結合的CRM蛋白的表達是優選的。用於表達和生產CRM蛋白的優選的表達系統包括具有細胞內氧化狀態的微生物。優選的表達系統可以是重組或天然的真核或原核細胞,其中重組細胞包括包含非天然CRM編碼序列的細胞。優選的原核細胞是大腸桿菌的菌株或包含一個或多個遺傳改變(例如一個或多個缺失或突變)的另一細菌菌株。優選地,與野生型相比,一個或多個遺傳改變將細胞的細胞質的氧化還原狀態改變為更加氧化的狀態,例如公開於美國專利號7,410,788(通過引用結合到本文中)。改變優選地降低一種或多種二硫化物還原酶基因和/或其它基因的活性,所述其它基因降低細胞質的氧化狀態。優選地,活性降低歸因於一種、兩種或多種二硫化物還原酶或其它基因的無表達或表達降低,或降低一種或多種表達的二硫化物還原酶蛋白或其它蛋白的活性的一種或多種突變。優選的微生物細胞(例如,重組、改造或天然的真核或原核細胞)菌株生產仍為功能蛋白的包含二硫鍵的天然摺疊蛋白的能力增加。本發明的方法生產一定量的包含完全、截短或修飾的CRM胺基酸序列的CRM蛋白。根據本發明生產的CRM蛋白的量是令人驚訝的,例如600mg或更多的CRM蛋白/升細菌細胞培養物。本發明的一個實施方案涉及生產大量CRM蛋白和優選地CRM197的方法。生產量通常按mg/L的細菌細胞培養物量化。根據本發明方法的CRM蛋白生產是200mg/L或更多、300mg/L或更多、400mg/L或更多、500mg/L或更多、600mg/L或更多、700mg/L或更多、800mg/L或更多、900mg/L或更多、1,000mg/L或更多、1,500mg/L或更多、或2,000mg/L或更多。本發明優選量的CRM197包括包含全長和截短的CRM蛋白以及修飾的CRM蛋白胺基酸序列的相關蛋白。修飾包括一個或多個保守胺基酸缺失、取代和/或添加。保守修飾是保持分子的功能活性和/或免疫原性的修飾,但可增加或降低活性和/或免疫原性。CRM的保守修飾的實例包括但不限於胺基酸修飾(例如,單個、兩個和另外的短胺基酸添加、缺失和/或取代)、可容易用於綴合形成疫苗的39個賴氨酸殘基的α-氨基(賴氨酸的伯胺基)之外的修飾、由於DT的血清型變化導致的修飾、增加免疫原性或增加綴合效率的修飾、基本上不改變與肝素的結合的修飾、保留正確摺疊或三維結構的修飾、和/或不顯著改變提供對DT的保護性免疫的蛋白或蛋白部分的免疫原性的修飾。用於本發明方法的重組細胞優選地是大腸桿菌細菌,和優選地,經遺傳改造以改變細胞質的氧化還原狀態至更加氧化的狀態的大腸桿菌(例如通過一種或多種二硫化物還原酶基因的突變,例如氧化還原酶、二氫葉酸還原酶、硫氧還蛋白還原酶、穀氨酸半胱氨酸裂解酶、二硫化物還原酶、蛋白還原酶和/或穀胱甘肽還原酶)。優選地,一種或多種二硫化物還原酶基因被突變,和賦予無功能性或少量功能性,使得與野生型相比,細胞的細胞質的氧化還原狀態被改變至更加氧化的狀態。氧化的蛋白摺疊涉及二硫橋的形成和異構化,並在許多蛋白包括CRM197的穩定性和溶解性方面起關鍵作用。二硫橋的形成和斷裂通常通過硫醇-二硫化物氧化還原酶催化。這些酶的特徵為一個或多個Trx摺疊,其由通過三個α-螺旋圍繞的四鏈β−片組成,具有CXXC氧化還原活性位點基序。各種Trx組件的裝配已經用於構建在原核和真核生物中存在的不同的硫醇氧化還原酶。在細菌周質中,通過DsbB-DsbA和DsbD-DsbC/DsbE/DsbG對的組合作用,蛋白保持在合適的氧化狀態(Inaba2009,Gruber等2006)。許多蛋白表達系統是本領域眾所周知的,和市售可得的。特別優選的微生物包括大腸桿菌表達菌株,例如化學感受態的大腸桿菌K12細胞,其經工程改造以在細胞質中形成二硫鍵鍵合的蛋白(例如,ORIGAMI™(EMDMillipore)和SHUFFLE™(NewEnglandBiolabs))。也可使用氧化型氧化還原狀態提高的其它菌株和細胞類型和其它大腸桿菌菌株。例如,ORIGAMI™2宿主菌株是在硫氧還蛋白還原酶(trxB)和穀胱甘肽還原酶(gor)兩個基因中具有突變的K-12衍生菌,其極大提高在大腸桿菌細胞質中的二硫鍵形成。這些菌株是卡那黴素敏感性的;像原始的Origami菌株一樣,仍通過四環素選擇gor突變。為了減少在分子之間形成二硫鍵的可能性,建議包含trxB和gor突變的菌株僅用於表達為了正確摺疊而需要二硫鍵形成的蛋白。SHUFFLE™細胞是化學感受態大腸桿菌K12細胞,其經工程改造以在細胞質中形成包含二硫鍵的蛋白。優選地,這些細胞包含trxB和gor突變和細胞質蛋白伴侶二硫鍵異構酶DsbC(fhuA2[lon]ompTahpCgalλatt::pNEB3-r1-cDsbC(SpecR,lacIq)ΔtrxBsulA11R(mcr-73::miniTn10--TetS)2[dcm]R(zgb-210::Tn10--TetS)endA1Δgor∆(mcrC-mrr)114::IS10)。還優選地,細胞適合於T7啟動子驅動的蛋白表達和具有基因型F´lac,pro,lacIQ/Δ(ara-leu)7697araD139fhuA2lacZ::T7gene1Δ(phoA)PvuIIphoRahpC*galE(orU)galKλatt::pNEB3-r1-cDsbC(SpecR、lacIq)ΔtrxBrpsL150(StrR)ΔgorΔ(malF)3。SHUFFLE™菌株組成型表達二硫鍵異構酶DsbC的染色體拷貝。DsbC促進校正錯誤氧化的蛋白為它們的正確形式。細胞質DsbC還是可有助於不需要二硫鍵的蛋白摺疊的蛋白伴侶。細菌培養物優選地在使得與較高的溫度(例如37°C)下的溶解性相比表達的蛋白的溶解性增加的溫度下培養(例如CRM或CRM197)。優選的培養溫度是30°C或更低、優選地25°C或更低、優選地20°C或更低、優選地18°C或更低和優選地在15°C和32°C之間。本發明的另一實施方案涉及用於生產CRM的載體和生產整個CRM蛋白或其一部分(例如優選地CRM197)的方法,所述CRM蛋白在細胞和優選地原核細胞的細胞質中可溶。以前在大腸桿菌中在細胞內表達CRM的嘗試基於僅編碼CRM序列的單順反子mRNA,和導致包涵體形成。在細胞的細胞質中生產可溶性CRM的方法使用提供在多順反子mRNA中的CRM的轉錄的表達載體開發。多順反子mRNA是指編碼兩種或更多種多肽的信使RNA。在原核細胞中,參與相同生物化學或生理學途徑的基因往往歸類為操縱子,其控制基因轉錄為單一的多順反子mRNA。操縱子中的基因(順反子)受核糖體結合位點序列的控制,和可通過多個核苷酸或甚至重疊序列分開。例如,第一個基因的終止密碼子在第二個基因的起始密碼子的下遊,如在半乳糖操縱子中。已經表明在操縱子中的基因位置通過翻譯和mRNA穩定性作用也強烈影響基因表達水平(Smolke,C.D.和Keasling,J.D.(2002)Effectofgenelocation,mRNAsecondarystructures,andRNasesitesonexpressionoftwogenesinanengineeredoperon.Biotechnol.Bioeng.80,762−76)。發現下遊基因表達水平經過翻譯偶聯,通過上遊基因表達而提高(Schümperli,D.,McKenney,K.,Sobieski,D.a和Rosenberg,M.(1982)TranslationalcouplingatanintercistronicboundaryoftheEscherichiacoligalactoseoperon.Cell30,865−71)。本發明的一個優選的實施方案是包含原核啟動子和兩個編碼多肽(其中之一是CRM)的順反子的載體。各個順反子包含核糖體結合位點和起始密碼子,例如ATG。本發明還包括誘導表達載體生產CRM蛋白和分離表達的CRM蛋白。在一個優選的實施方案中,CRM序列之前的第一個順反子包含與CRM順反子重疊的T7標籤序列,使得第一個順反子的終止密碼子在CRM的起始密碼子的下遊(例如SEQIDNO15)。表達增強子按SEQIDNO16或SEQIDNO17進一步修飾。CRM編碼序列之前的第一個順反子被稱為「表達增強子序列」(EES)。表達載體包含(1)啟動子,接著是核糖體結合位點和表達增強子序列,和(2)核糖體結合位點和ATG密碼子和CRM編碼序列。重組細胞可以是原核或真核細胞。優選地,重組細胞是原核細胞,例如大腸桿菌細胞或大腸桿菌的衍生菌或菌株。優選地,重組細胞修飾包含降低活性的一種或多種二硫化物還原酶,例如氧化還原酶、二氫葉酸還原酶、硫氧還蛋白和硫氧還蛋白還原酶、蛋白還原酶或穀胱甘肽還原酶的一種或多種。優選地,與非-重組細胞相比,降低活性的一種或多種二硫化物還原酶改變重組細胞的細胞質的氧化還原狀態至氧化狀態。優選地,CRM編碼序列編碼一個或多個CRM表位、CRM肽序列、CRM結構域或其組合。優選地,由細胞表達的CRM蛋白是可溶性的,和是細胞內的、周質的或分泌的。優選地,重組細胞在約15°C至約32°C的溫度下增殖。本發明的另一實施方案包括包含可表達的CRM序列和優選地CRM197的序列的重組細胞,例如細菌、哺乳動物或昆蟲細胞。優選的宿主細胞包括但不限於經遺傳改造以改變細胞質的氧化還原狀態至更加氧化的狀態的細胞。優選的細胞包括原核或真核細胞,例如大腸桿菌細胞表達系統,杆狀病毒表達系統和其它細菌和/或真核細胞表達系統。優選地,細胞包含用於表達外源或非天然序列例如CRM肽的蛋白表達系統。還優選的,待表達的序列由表達載體構成,其包含一個或多個誘導型啟動子(例如,自動誘導型,優選地用特定培養基)、起始密碼子(例如ATG)、核糖體結合位點、非特定的多肽序列和轉錄為多順反子mRNA的CRM編碼序列。還優選地,表達載體在核糖體結合位點和ATG起始密碼子之間或在起始密碼子和待表達的序列之間包含修飾的序列。優選的修飾序列或間隔基序列包括例如,多於9個或少於9個的多個核苷酸(例如7-12個核苷酸),和優選地不是9個核苷酸。可用於表達系統的間隔基核苷酸的具體實例包括但不限於GATATAC(SEQIDNO3)、GATATACCA(SEQIDNO4)和GATATACCATAT(SEQIDNO5)。因此,本發明的另一實施方案包含CRM的表達構建體、核苷酸和胺基酸序列,含或不含定義的間隔基序列並且含和不含宿主細胞。本發明的另一實施方案涉及重組CRM197蛋白以及重組CRM在大腸桿菌或另一宿主細胞中使用具有誘導型啟動子和/或在核糖體結合位點和ATG起始密碼子之間的修飾序列的表達載體的表達。優選地,表達載體包括乳糖/IPTG誘導型啟動子,優選地tac啟動子和在核糖體結合位點和ATG起始密碼子之間的序列。優選地,表達系統在起始密碼子和表達序列之間包含間隔基,其由多於9個或少於9個的多個核苷酸構成(例如7-12個核苷酸),和優選地不是9個核苷酸。可用於表達系統的間隔基核苷酸的具體實例包括但不限於本文鑑定的那些。令人驚訝的發現,與9個核苷酸的間隔基相比,使用長度為7或12的間隔基導致顯著增加CRM197表達水平。本發明的另一實施方案包括CRM的表達構建體、核苷酸和胺基酸序列,含或不含本文所公開的定義的間隔基序列和含或不含增強子區。增強子區通過增強CRM的正確摺疊的翻譯偶聯促進下遊CRM序列的表達,導致蛋白可溶性。增強子區還通過添加促進核酸識別以增加表達的一個或多個序列,促進蛋白表達(例如,起始密碼子、酶結合位點、翻譯或轉錄因子結合位點)。優選地,本發明的增強子包含核糖體結合位點與編碼序列上遊的起始密碼子和編碼序列,所述編碼序列不同於CRM蛋白的編碼序列。本發明的另一實施方案涉及根據本發明的方法純化的重組CRM,和具體而言CRM197。純化優選地包括肝素或肝素-樣親和色譜法。令人驚訝的發現,CRM197包含典型的肝素結合位點XBBXBX(SEQIDNO6)的基於序列的基序,其中B是賴氨酸或精氨酸和X是親水殘基(CardinAD,WeintraubHJ.,1989:Molecularmodelingofprotein-glycosaminoglycaninteractions.Arteriosclerosis9:21–32)。該基序位於CRM197受體-結合結構域中並包含以下胺基酸:GRKIRMRCR(SEQIDNO7),其中G(甘氨酸)、I(異亮氨酸)、M(甲硫氨酸)和C(半胱氨酸)是親水殘基。肝素結合位點的存在允許在純化中使用肝素或肝素-樣樹脂。肝素-樣樹脂包括包含硫酸官能團的樹脂,例如硫酸葡聚糖,例如硫酸葡聚糖(Sterogene)、CaptoDevirs(GE)或硫酸酯,例如CellufineSulfate(AsahiKaseiBioprocess)。在第一步中,粗大腸桿菌提取物可例如優選地通過離心或深過濾澄清。任選澄清的裂解物可優選地通過加入對蛋白溶解度有影響的鹽和鹽析CRM197進一步分級。在第二步中,澄清的裂解物或包含CRM197的再溶解的鹽析部分可在當CRM197流動通過的條件下施加至例如陰離子交換樹脂。在第三步中,包含CRM197的流動通過部分可施加至柱。優選的柱樹脂包括但不限於硫酸葡聚糖樹脂、CELLUFINE™樹脂(ChissoCorporation;色譜凝膠)、活性硫酸化樹脂、磷酸鹽樹脂或肝素或肝素-樣樹脂。優選地,CRM與樹脂結合在低鹽緩衝液中進行和在較高鹽緩衝液中洗脫,得到高度純化的CRM197。優選的結合緩衝液包含例如一種或多種離液劑、NaCl、KCl、甘油、異丙醇、乙醇、精氨酸、乙酸鹽、胍、尿素、ATP、一種或多種一-、二-、三-和/或多-磷酸鹽、硫酸鹽或焦磷酸鹽,和其組合。優選的洗脫緩衝液包含例如,較高濃度的一種或多種結合緩衝液組分。其它優選的純化方法包括陰離子交換色譜法、疏水相互作用色譜法和/或Cibacron-Blue樹脂(CN101265288A、美國專利號8,383,783)的任何一種或組合。本發明的純化方法以如本文所述的高收率和大於80%、優選地大於85%、優選地大於90%、優選地大於95%、優選地大於99%純度水平和優選地甚至更高的純度生產重組CRM蛋白(例如CRM197)。本發明的另一實施方案涉及表徵包含受體結合結構域(見SEQIDNO2)的重組DT和CRM蛋白(例如結合活性),和具體而言CRM197的方法。這些方法包括測定包含DT受體結合結構域的天然或修飾序列的蛋白的結合活性。這樣的修飾優選地保留CRM結合HB-EGF(肝素結合表位生長因子)的能力。所述方法可適用於粗製和純化的CRM197兩者。結合活性代表了可溶形式的白喉毒素受體HB-EGF(DTR)的結合。這些方法優選地包括測定CRM197與DTR的結合和用對正確摺疊的結構、複合物、結合和/或結合位點有特異性的分子(例如抗體、抗體片段、抗原)和優選地以ELISA形式檢測。測定和定量結合的測定法允許快速檢測CRM197是正確摺疊的,因為僅正確摺疊的CRM197結合受體。因此,所述方法監測在開發、生產和純化過程中製備的CRM197和相關蛋白的正確摺疊。此外,該表徵方法可用於在與另一分子綴合後鑑定和跟蹤CRM蛋白,例如在疫苗生產中。使用本發明的檢測方法,在開發用於治療和/或預防患者的疾病和病症的疫苗過程中可監測正確摺疊和構造的綴合的CRM蛋白。本發明的另一實施方案包括用於綴合CRM蛋白的方法,用於疫苗生產,例如通過與多糖綴合。還包括蛋白、肽、寡糖和半抗原的綴合。本發明的另一實施方案包括包含本發明的CRM蛋白的疫苗。本發明的另一實施方案涉及與另一分子例如另一蛋白或多糖經基因或化學融合的本發明的CRM蛋白。融合優選地通過在分子之間的一個或多個共價鍵進行。以下實施例舉例說明了本發明的實施方案,但不應視為限制本發明的範圍。實施例實施例1.自在RBS和起始ATG密碼子之間包含7、8、9或12個核苷酸序列的表達載體檢測的CRM197表達將編碼成熟CRM197的DNA以多順反子形式克隆至表達載體,產生以下DNA調節和編碼片段的序列:tac啟動子-核糖體結合位點-ATG密碼子-T7標籤-核糖體結合位點-ATG密碼子-CRM編碼序列-終止密碼子。不同的大腸桿菌菌株,包括ORIGAMI2、C41,作為表達菌株進行測試(圖1)。實施例2.CRM197在允許在細胞質中形成二硫鍵的Origami2大腸桿菌表達菌株中可溶性表達CRM197在37°C不溶性表達。當表達溫度下降至低於37℃時,在ORIGAMI™2細胞和SHUFFLE™細胞中而非在其它測試的大腸桿菌菌株中表達的蛋白的溶解度增加。當在ORIGAMI™2細胞中在18°C下表達時,CRM197大部分是可溶的。實施例3.在CRM197表達中的表達增強子序列(EES)EES在包含CRM的載體中促進CRM序列的轉錄和產生多順反子mRNA,其翻譯成兩種蛋白,短的EES肽和CRM肽。對於潛在的3D結構形成,分析天然CRM基因的編碼序列,和發現包含多個潛在的髮夾結構,其可抑制翻譯。創建CRM序列,其將可能產生沒有髮夾結構但仍翻譯相同的CRM胺基酸序列的mRNA。該基因序列稱為最佳CRM序列和包含SEQIDNO8。最佳CRM序列在大腸桿菌(例如BL21)和經改造包含氧化細胞質的大腸桿菌(例如Shuffle)中均良好表達。自多順反子mRNA翻譯的CRM肽產生全長蛋白,和認為其比天然CRM編碼序列更穩定。不像天然CRM序列,最佳CRM序列作為全長和作為可溶性蛋白在Shuffle細胞中表達。此外,與天然CRM相比,以較低的細胞密度和以與色譜樹脂的結合增加觀察到最佳CRM序列的更高表達,使CRM蛋白的生產水平更高。實施例4.硫酸銨沉澱來自細胞裂解物的CRM197表達CRM197的SHUFFLE™細胞使用微流化儀打開,向細胞裂解物中加入1M的氯化鈉。向其中加入足夠的硫酸銨至等於1M,接著以20,000xg離心30分鐘,這除去錯誤摺疊的CRM197和大部分的細菌蛋白。在澄清後,硫酸銨濃度進一步增加至2.2M。收集主要是CRM197的沉澱物和再溶於低電導率緩衝液。實施例5.在肝素柱上CRM197的純化將硫酸銨沉澱的CRM197再溶於20mMTris-HClpH8.0,達到電導率5mS/cm和上樣到包含肝素瓊脂糖CL-6B樹脂(GE)的柱上。在以下條件下進行純化:流速5ml/min,洗滌緩衝液A:20mMTris-HClpH8。用緩衝液B0-100%梯度,緩衝液B:緩衝液A+1MNaCl以20個CV進行洗脫。洗脫的CRM197通過SDS-PAGE在還原和非還原條件下分析。洗脫的CRM197的純度大於95%。用DTT還原的蛋白作為單一多肽顯示,證實了完整形式的CRM197在大腸桿菌中表達。實施例6.在CaptoDevirs柱上的CRM197的純化表達CRM197的SHUFFLETM細胞使用微流化儀在1xPBSpH7.4、1%焦磷酸鈉中打開。裂解物使用深過濾澄清。將澄清的裂解物上樣到包含QSepharoseXL(GE)的柱上,和收集流動通過的部分(flowthroughfraction)。為減少體積和電導率,將流動通過部分使用10K盒(cassette)(Sartorius)進行切向流過濾。CaptoDevirs樹脂用25mM焦磷酸鈉緩衝液pH8.0平衡。CRM197在以下條件下與柱結合:電導率低於10mS/cm,在包含離液劑(例如在該情況下為尿素)的結合緩衝液中,洗滌緩衝液為25mM焦磷酸鈉pH8.0。用NaCl進行洗脫。洗脫的CRM197通過SDS-PAGE在還原和非還原條件下分析。洗脫的CRM197的純度大於95%。用DTT還原的蛋白作為單一多肽顯示,證實了CRM197在純化過程期間保持完整。實施例7.用於CRM197表徵的結合測定將重組可溶性白喉毒素受體HB-EGF(DTR)(Sigma)結合至ELISA板。5%脫脂奶粉的封閉液用於防止高背景。將在1xPBSpH7.4、0.1%Twin20中稀釋的重組CRM197在37°C在板上孵育1小時。與HB-EGF結合的CRM197通過兔多克隆抗-CRM197抗體和與大豆過氧化物酶綴合的山羊抗兔抗體(FinaBioSolutions;Rockville,MD)檢測。變性的重組CRM197不與受體結合。實施例8.在大腸桿菌中生產的CRM197結合DTR,其類似於來自棒狀桿菌和假單胞菌的CRMELISA板用可溶性HB-EGF(肝素-結合EGF-樣生長因子)包被,和用5%脫脂奶粉封閉。CRM197與受體結合,和用兔抗-CRM197多克隆抗體和與SBP綴合的山羊抗-兔多克隆抗體檢測。在大腸桿菌中表達的CRM197顯示對HB-EGF的親和力與在棒狀桿菌和假單胞菌中生產的CRM相同。實施例9CRM197是載體蛋白CRM197按照本發明的方法表達和純化(實施例1)和使用CDAP化學,化學連接(綴合)至肺炎球菌莢膜多糖血清型14和6B(Lees,A.Producingimmunogenicconstructsusingsolublecarbohydratesactivatedviaorganiccyanylatingreagents.見美國專利號5,651,971;5,693,326和5,849,301)。綴合物自未綴合的蛋白和多糖純化。根據表1的時間表用綴合物皮下免疫BALB/c雌性小鼠。在完全弗氏佐劑中免疫小鼠和在不完全弗氏佐劑中加強兩次,並在第57天採血。表1血清型在CFA中初次*加強IFA**加強IFA**D576B20ug10ug,第28天10ug,第48天採血1420ug5ug,第21天5ug,第48天採血*60%完全弗氏佐劑;**60%不完全弗氏佐劑通過ELISA在用2µg/ml的Pn6B或Pn14(來自ATCC)包被的Brandtech免疫級板上使用γ-特異性檢測,測試血清的反應性。圖2的結果表明與Pn6B的強反應性。小鼠5086用於雜交瘤生產,並且得到的雜交瘤中的三個用於製備高特異性小鼠抗-6B單克隆抗體。針對Pn14包被板的血清滴定的結果顯示在圖3中。小鼠1397隨後用於生產與P14多糖有反應的四種高特異性小鼠單克隆抗體。未綴合的多糖不產生顯著的ELISA吸光度。考慮本文公開的本發明的說明書和實踐,本發明的其它實施方案和用途對本領域技術人員而言將是顯然的。本文引用的所有參考文獻,包括所有出版物、美國和其它國家的專利和專利申請,明確和完整地通過引用結合到本文中。術語「包含」,在任何時候使用時,意圖包括術語「由……組成」和「基本上由……組成」。此外,術語「包含」、「包括」、「含有」等不意圖限制。預期說明書和實施例僅認為是示例性的,本發明的真實範圍和精神由隨附權利要求書表明。序列SEQIDNO1 CRM197GADDVVDSSKSFVMENFSSYHGTKPGYVDSIQKGIQKPKSGTQGNYDDDWKEFYSTDNKYDAAGYSVDNENPLSGKAGGVVKVTYPGLTKVLALKVDNAETIKKELGLSLTEPLMEQVGTEEFIKRFGDGASRVVLSLPFAEGSSSVEYINNWEQAKALSVELEINFETRGKRGQDAMYEYMAQACAGNRVRRSVGSSLSCINLDWDVIRDKTKTKIESLKEHGPIKNKMSESPNKTVSEEKAKQYLEEFHQTALEHPELSELKTVTGTNPVFAGANYAAWAVNVAQVIDSETADNLEKTTAALSILPGIGSVMGIADGAVHHNTEEIVAQSIALSSLMVAQAIPLVGELVDIGFAAYNFVESIINLFQVVHNSYNRPAYSPGHKTQPFLHDGYAVSWNTVEDSIIRTGFQGESGHDIKITAENTPLPIAGVLLPTIPGKLDVNKSKTHISVNGRKIRMRCRAIDGDVTFCRPKSPVYVGNGVHANLHVAFHRSSSEKIHSNEISSDSIGVLGYQKTVDHTKVNSKLSLFFEIKSSEQIDNO2 CRM197的結構域SPGHKTQPFLHDGYAVSWNTVEDSIIRTGFQGESGHDIKITAENTPLPIAGVLLPTIPGKLDVNKSKTHISVNGRKIRMRCRAIDGDVTFCRPKSPVYVGNGVHANLHVAFHRSSSEKIHSNEISSDSIGVLGYQKTVDHTKVNSKLSLFFEIKSSEQIDNO3 GATATAC 間隔基SEQIDNO4 GATATACCA 間隔基SEQIDNO5 GATATACCATAT 間隔基SEQIDNO6 XBBXBX 推定的肝素結合位點SEQIDNO7 GRKIRMRCR 肝素結合位點SEQIDNO8 最佳CRM序列ATGGGTGCTGATGATGTTGTTGATTCCTCTAAGTCTTTCGTGATGGAAAATTTCTCGTCCTATCACGGTACCAAGCCTGGCTATGTGGATAGCATTCAAAAGGGTATTCAAAAACCGAAGTCTGGTACCCAGGGCAACTACGATGACGATTGGAAAGAGTTTTACAGCACCGACAACAAATATGACGCGGCAGGCTACAGCGTTGATAATGAAAATCCGCTGAGCGGTAAGGCTGGCGGCGTCGTTAAGGTTACCTATCCGGGTCTGACGAAAGTGCTGGCCCTGAAAGTTGACAATGCTGAAACCATCAAAAAAGAACTGGGTCTGAGCTTGACCGAGCCGCTGATGGAACAGGTTGGTACTGAAGAATTCATTAAACGTTTTGGTGACGGCGCGAGCCGTGTTGTGCTGTCCCTGCCGTTTGCCGAGGGTTCTAGCTCCGTGGAGTATATCAACAATTGGGAACAGGCGAAAGCGTTGAGCGTCGAGCTGGAAATCAATTTCGAGACTCGTGGTAAGCGTGGCCAAGATGCGATGTACGAGTACATGGCCCAGGCATGTGCGGGTAACCGCGTCCGTCGCAGCGTCGGCAGCTCCCTGAGCTGCATTAACCTGGACTGGGACGTGATCCGCGACAAGACTAAGACCAAGATTGAGAGCCTGAAAGAGCACGGTCCGATTAAGAACAAAATGTCCGAGTCTCCGAACAAAACGGTGAGCGAAGAAAAAGCCAAACAGTATCTGGAAGAATTCCATCAGACCGCCCTGGAGCACCCAGAGCTGAGCGAGCTGAAAACCGTCACCGGCACGAATCCGGTTTTTGCGGGTGCGAACTACGCGGCATGGGCAGTCAATGTTGCGCAAGTCATCGACAGCGAAACGGCTGATAACTTGGAGAAAACCACCGCGGCACTGAGCATTCTGCCGGGCATCGGTAGCGTTATGGGCATTGCGGACGGTGCCGTGCATCACAATACCGAAGAAATTGTCGCGCAGAGCATCGCATTGTCTAGCCTGATGGTTGCACAGGCCATTCCGCTGGTAGGCGAATTGGTGGATATCGGTTTCGCGGCTTACAATTTCGTTGAGTCGATCATTAACCTGTTTCAAGTCGTTCACAATAGCTATAACCGTCCGGCATACAGCCCGGGTCATAAGACGCAACCGTTTCTGCATGATGGCTATGCCGTGAGCTGGAACACGGTCGAGGATTCGATTATCCGTACCGGTTTTCAGGGTGAGAGCGGTCACGACATCAAAATCACCGCGGAGAACACGCCGCTGCCTATTGCGGGCGTCCTGCTGCCGACGATCCCGGGCAAACTGGACGTTAACAAGAGCAAGACCCATATCAGCGTCAACGGTCGTAAGATTCGCATGCGTTGTCGTGCAATCGACGGTGACGTGACGTTCTGCCGCCCAAAAAGCCCGGTGTACGTGGGTAACGGCGTGCACGCGAATCTGCATGTCGCGTTCCACCGCTCCTCAAGCGAGAAAATCCACAGCAATGAAATTAGCAGCGACAGCATTGGTGTGTTGGGCTACCAAAAGACCGTGGATCACACCAAGGTTAATAGCAAGCTGAGCCTGTTCTTTGAGATCAAAAGCSEQIDNO9 非最佳CRM序列ATGGGTGCCGATGACGTGGTTGACTCTTCCAAAAGCTTCGTCATGGAAAACTTCAGCTCCTATCACGGCACTAAACCGGGTTATGTCGACAGCATCCAGAAAGGCATCCAGAAACCGAAATCTGGCACTCAGGGTAACTATGACGACGACTGGAAAGAGTTCTACTCTACCGACAACAAATACGACGCGGCTGGTTATTCTGTGGACAACGAAAACCCGCTGTCTGGTAAAGCTGGTGGTGTTGTTAAAGTGACCTACCCGGGTCTGACCAAAGTTCTGGCTCTGAAAGTGGACAACGCCGAAACCATCAAAAAAGAACTGGGTCTGTCTCTGACCGAACCGCTGATGGAACAGGTAGGTACCGAGGAATTCATCAAACGTTTTGGTGATGGTGCGTCCCGTGTTGTACTGTCTCTGCCATTTGCCGAAGGTTCTAGCTCTGTCGAGTACATCAACAACTGGGAGCAGGCCAAAGCTCTGTCTGTGGAACTGGAAATCAACTTCGAGACCCGTGGTAAACGTGGTCAGGACGCAATGTATGAATACATGGCACAGGCTTGCGCGGGTAACCGTGTACGTCGTTCTGTAGGTTCTTCCCTGTCTTGCATCAACCTGGACTGGGATGTCATCCGTGACAAAACCAAAACCAAAATCGAGTCCCTGAAAGAGCACGGTCCGATCAAAAACAAAATGAGCGAATCTCCGAACAAAACGGTCTCTGAGGAAAAAGCGAAACAGTACCTGGAAGAATTCCATCAGACCGCCCTGGAACACCCGGAACTGTCTGAACTGAAAACCGTTACCGGTACTAACCCGGTTTTCGCAGGTGCTAACTACGCAGCGTGGGCGGTTAACGTAGCCCAGGTAATCGATTCCGAAACCGCAGACAACCTGGAAAAAACGACTGCGGCTCTGTCTATTCTGCCGGGTATTGGTAGCGTGATGGGTATTGCAGATGGTGCAGTTCACCACAACACGGAAGAAATCGTTGCGCAGTCTATCGCTCTGTCTTCTCTGATGGTAGCACAGGCGATCCCGCTGGTTGGTGAACTGGTTGACATTGGCTTCGCGGCCTACAACTTCGTTGAATCCATCATCAACCTGTTCCAGGTTGTGCACAACTCTTACAACCGTCCAGCTTACTCTCCGGGTCACAAAACCCAGCCGTTCCTGCACGACGGTTATGCGGTTTCTTGGAACACCGTTGAAGACAGCATCATCCGTACTGGTTTCCAGGGTGAATCTGGCCACGACATCAAAATCACTGCTGAAAACACCCCGCTGCCGATCGCAGGTGTTCTCCTGCCAACTATTCCGGGTAAACTGGACGTGAACAAATCCAAAACGCACATCTCCGTGAACGGTCGTAAAATCCGCATGCGTTGTCGTGCGATTGATGGTGACGTTACTTTCTGTCGTCCGAAATCTCCGGTCTACGTAGGTAACGGTGTACATGCTAACCTCCATGTAGCGTTCCACCGTTCTTCTTCCGAGAAAATCCACTCCAACGAGATCTCTAGCGACTCTATCGGTGTTCTGGGTTACCAGAAAACCGTTGACCACACCAAAGTGAACTCCAAACTCAGCCTGTTCTTCGAAATCAAATCTSEQIDNO10 crm7GAGCTCTAAGAAGGAGATATACATGGGTGCCGATGACGTGGTTGACTCTSEQIDNO11 crm7_2GAGCTCTTAAGAAGGAGATATACATGGGTGCCGATGACGTGGTTGACTCTSEQIDNO12 crm8GAGCTCTAAGAAGGAGATATACAATGGGTGCCGATGACGTGGTTGACTCTSEQIDNO13 crm9GAGCTCTAAGAAGGAGATATACACATGGGTGCCGATGACGTGGTTGACTCTSEQIDNO14 crm12GAGCTCTAAGAAGGAGATATACCATATATGGGTGCCGATGACGTGGTTGACTCTSEQIDNO15TCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGGCTAGCATGACTGGTGGACAGCAAATGGGTCGGGATCCGAATTCGAGCTCTAAGAAGGAGATATACCSEQIDNO16TCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGGCTAGCATGACTGGTAAGGAGATATACCSEQIDNO17TCTAGAAATAATTTTGTTTAACTTTAAGAAGGAGATATACATATGGCTAGCATGACTGGTGCGMAYCCATTCAGTGAAGAAGRAGSTTYATTT當前第1頁1 2 3