在噬菌體顆粒上展示環肽的新方法與流程
2023-08-03 16:26:26 7
發明領域
本發明涉及在噬菌體外被蛋白的c-端處在噬菌體顆粒的表面上展示環肽的方法。
背景技術:
環肽是採取環狀環結構的多肽,已知其具有多種生物活性,例如抗菌活性、免疫抑制活性或抗腫瘤活性。若干種在天然界發現的環肽被用於臨床,例如抗菌劑短桿菌肽s、短桿菌酪肽和萬古黴素,或者具有免疫抑制活性的環孢黴素a。受具有生物活性的天然環肽的啟發,已經著力於用遺傳和合成方法開發人工環肽。
一類新興的具有環狀結構的生物分子是核糖體合成的肽,其要求廣泛的翻譯後修飾來形成生物活性肽。大部分核糖體合成的天然肽被翻譯成由前導肽和核心肽組成的前體。前導用作識別序列,並募集酶體系(enzymaticmachinery),以在核心肽的特定殘基處安裝翻譯後修飾(ptm)。
因此,翻譯後修飾例如雜環化或大環化、脫水、乙醯化、糖基化、滷化、異戊烯化和差向異構化,不僅使這些肽形成生物活性,而且直接有助於該類肽的許多代表性成員中發現的優異穩定性,因此使其成為藥物開發有吸引力的候選物。
羊毛硫肽(lanthipeptides)和羊毛硫抗生素形成一類獨特的核糖體合成的翻譯後修飾抗生素肽,其由革蘭氏陽性細菌產生,並主要對其發揮作用(綜述參見knerr和vanderdonk,annu.rev.biochem.2012.81:479-505)。天然羊毛硫抗生素例如乳酸鏈球菌素(nisin)或枯草菌素(stbtilin)已經深入研究,並在商業上用於食品工業用於製造並存儲乳製品例如奶酪。
作為具有抗微生物活性的肽的子類,羊毛硫肽和羊毛硫抗生素含有由硫醚胺基酸羊毛硫氨酸(lan)和3-甲基羊毛硫氨酸(melan)形成的分子內硫醚-橋或環,其保護這些肽不受蛋白水解降解,並使其熱穩定性增加。硫醚-橋的安裝始於絲氨酸或蘇氨酸分別酶脫水成不飽和脫氫丙氨酸(dha)和脫氫氨基丁酸(dhb),之後進行半胱氨酸硫醇的分子內麥可加成,並通過羊毛硫肽合成酶介導(i類為lanb和lanc,ii類為lanm,iii類為lankc,iv類為lanl)。在i類羊毛硫肽中,絲氨酸/蘇氨酸脫水和隨後的環化分別通過lanb型脫水酶和lanc型環化酶進行,而在ii類羊毛硫肽中,單一的雙功能lanm型酶進行兩種反應。有趣的是,不飽和dha具有很高的化學反應性,可以在弱鹼性條件下很容易與半胱氨酸或賴氨酸的側基反應,分別產生非立體選擇性硫醚-橋和賴丙氨酸-橋。iii類和iv類羊毛硫肽的生物合成分別由多功能lankc和lanl型酶支持,其特徵在於氨基端的磷-ser/磷-thr裂解酶結構域、中心的類似於激酶的結構域和羧基端的類似於lanc的結構域(環化酶)(vanderdon等人2014currentopinioninstructuralbiology2014,29:58-66)。
過去數年間,羊毛硫肽生物合成的原理越來越多地適應於具有環狀結構的人工生物活性肽的發現和產生。
在2004年首次提出羊毛硫肽合成酶可以有利地用於將ptm(例如硫醚-橋)引入到正常未修飾的肽中,以提高肽的穩定性和/或改變其活性(kuipers等人2004.j.biol.chem.279,22176-22182)。在wo2006/062398中,已經顯示所關注的肽可以在宿主細胞中通過並非常規羊毛硫抗生素酶複合物一部分的分離的羊毛硫抗生素脫水酶(例如lanb)得以脫水。經進一步說明,經修飾的含有硫醚-橋的肽可以被其天然宿主中除專用羊毛硫抗生素轉運蛋白以外的蛋白質輸出系統分泌。
後來,在wo2012/005578中,說明含有硫醚-橋的多肽可以很容易地由表達羊毛硫抗生素的生物合成和輸出機制的宿主細胞(例如乳酸鏈球菌lactococcuslactis)的表面產生,並在其上展示。
更具體地,wo2012/005578提供了一種表達載體,其編碼包括n-端羊毛硫抗生素前導序列、受關注的要被翻譯後修飾成含有脫氫殘基或硫醚的多肽和c-端帶電膜錨定結構域的融合肽。另外提出了篩選具有所需活性的環肽的展示文庫。但是,僅能夠在天然地能夠產生羊毛硫抗生素的革蘭氏陽性宿主細胞特別是乳酸細菌上展示。
其他本領域已知的展示系統例如要求蛋白質輸出機制不同的革蘭氏陰性細菌的噬菌體展示並不視作是可供選擇者,因此在現有技術中不可行。
噬菌體展示的歷史基於以下論證始於1985年:絲狀噬菌體耐受插入到其基因iii蛋白質(piii)中的外來蛋白質片段,並且還在噬菌體表面上呈現蛋白質片段(smith,1985)。ladner將該概念擴展至噬菌體表面上展示的(多)肽和/或蛋白質所有組成成分(repertoires)的篩選(wo1988/06630;wo1990/02809),自此以後,噬菌體展示經歷了迅猛發展,並導致大量成就。已經開發出許多種構建並篩選(多)肽/蛋白質噬菌體-展示庫的形式,大量的綜述性文章和專著涉及並總結了這些發展(例如,kay等人,1996;dunn,1996;mcgregor,1996)。為了將肽或蛋白質錨定在絲狀噬菌體表面上,主要使用與噬菌體外被蛋白的基因融合。優選與基因iii蛋白質(parmley&smith,1988)或其片段(bass等人,1990)和基因viii蛋白質(greenwood等人,1991)融合。在一情形中,使用基因vi(jespers等人,1995),最近,使用基因vii和基因ix的組合用於展示fv片段(gao等人,1999)。
迄今,僅有線性(多)肽和經過二硫鍵穩定化的環肽在噬菌體上得以成功展示(參見wo2000/077194、wo2009/098450)。最近地,在wo2012/019928中,在噬菌體上展示了與piii的n-端融合的microvividrink的線性前體。展示的線性前體的翻譯後修飾通過隨後將噬菌體與含有同源(cognate)修飾酶的細胞溶解產物一起孵育實現。但是,wo2012/019928沒有提供羊毛硫肽展示的充分公開,並且也沒有教導在噬菌體組裝之前經歷翻譯後修飾的肽的展示。
因此,對於將翻譯後修飾的環肽的展示從細菌翻譯為經典的噬菌體展示存在需求。
技術實現要素:
在現有技術中,含有硫醚-橋的肽的展示已經成功闡釋。但是,使用細菌用於展示伴隨有顯著缺點。例如,就電穿孔而言,革蘭氏陽性細菌例如乳酸乳球菌(l.lactis)的轉化效率降低,並具有凝集傾向,這使其難以處理並導致不穩定。因此,通過乳酸乳球菌展示的文庫多樣性不能超過超過~106,而典型的噬菌體展示文庫已知具有大於1012個不同的克隆。
因此,本公開內容的技術問題是要開發能夠使環肽在噬菌體顆粒上呈現的簡單、可靠的系統。通過提供特徵如本文所述的實施方式,實現該技術問題的解決方案。本公開內容將環肽與噬菌體外被蛋白的c-端連接,使得能夠在噬菌體顆粒上展示所述環肽。因此,c-端噬菌體展示顯示用於在pviii上(held等人,2004&weiss等人,2000)以及另外在piii外被蛋白上(fuh等人,2000)的線性肽,但沒有論及或提出環肽或經過噬菌體外被蛋白c-端翻譯後修飾的肽的展示。通常,常規的噬菌體展示使用噬菌體外被蛋白的n-端用於展示,並且沒有預計環肽可以僅通過噬菌體外被蛋白的c-端得以有效展示。
更具體地,本公開內容包括一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供帶有編碼前體環肽的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽的表達;
(c)使前體環肽內的一個或多個胺基酸殘基酶脫水;
(d)通過使一個或多個脫水的殘基與半胱氨酸或賴氨酸偶聯,形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
一方面,環肽包括通過將一個或多個脫水的殘基與半胱氨酸或賴氨酸偶聯形成的分子內鍵。另一方面,環肽包括通過將一個或多個脫水的殘基與半胱氨酸殘基或賴氨酸殘基偶聯形成的分子內鍵。另一方面,環肽包括通過將一個或多個脫水的胺基酸殘基與半胱氨酸或賴氨酸偶聯形成的分子內鍵。
因此,本公開內容使得可以在噬菌體上呈現環肽。本公開內容的技術方法,即將環肽與噬菌體外被蛋白的c-端連接在現有技術中沒有提供或提出。
因此,本公開內容涉及用於在噬菌體顆粒的表面上展示環肽的方法,其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。此外,本公開方法使得可以產生並篩選展示在噬菌體顆粒表面上的環肽的大的文庫。
在許多情形中,使用該方法呈現環肽是有利的。要求保護的展示包括環肽的文庫的方法具有實用性。
而且,包括通過使用要求保護的方法展示的環肽的文庫具有實用性。這樣的文庫包括範圍很廣的環肽,其可以針對所關注的靶標進行篩選。
使用所公開的方法使得可以在肽中基本上任意所需的位置處引入分子內鍵。特別關注的是具有生物活性的肽,例如意在用於治療用途的肽,因為引入一個或多個分子內鍵通常增加了肽的生物穩定性。而且,可以使用環結構改變肽的生物活性,例如,抗原特異性、受體結合親合力、抗微生物活性或酶特異性。例如,所關注的肽是激動性肽(agonisticpeptide)、拮抗性肽、醯胺化肽、激素、酶抑制劑、酶激活劑、受體配體、抑制性肽、羊毛硫抗生素蛋白質、病毒蛋白質、真核生物蛋白質、其突變體(例如,具體設計成可以在某一位點處進行修飾)、模擬物、同源物或與其相當的功能片段。該方法可以用於識別治療相關和治療活性的基於環肽的分子,或者可以用於表徵這些分子。
附圖說明
圖1:xa因子-切割報導基因檢驗證實了大腸桿菌(e.coli)中表達的環肽的修飾狀態
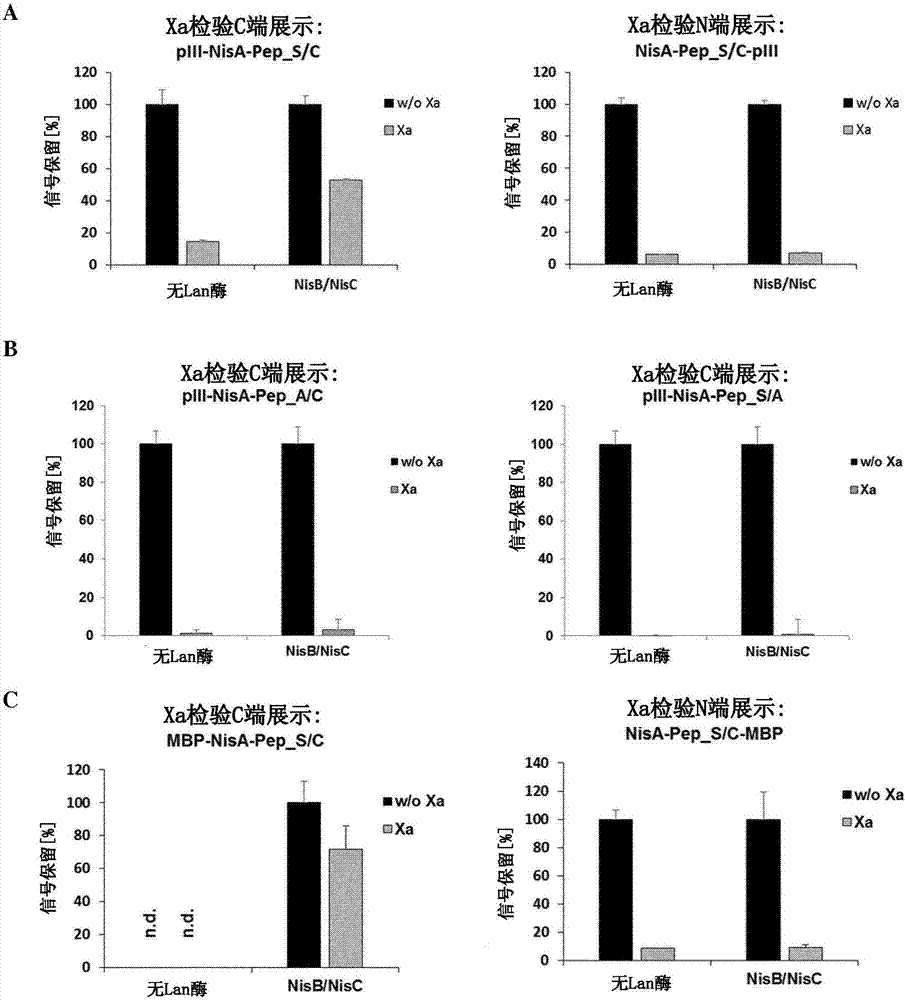
使用基於elisa的xa因子-切割報導基因檢驗,在存在或不存在修飾nisb/nisc酶的共表達的情況下,在大腸桿菌細胞溶解產物中評價帶有nisa-前導序列的可溶表達肽的修飾狀態。儘管在不存在nisb/nisc共表達的情況下(無lan-酶),產生的含有aswiegrwcn-基序的肽(seq-id.:1;加下劃線的xa因子識別序列)幾乎被xa因子完全切割,nisb/nisc的共表達導致酶促將硫醚-橋從脫氫丙氨酸(脫水絲氨酸)引入到半胱氨酸以及xa因子-切割抗性(左圖)。即使在nisb/nisc共表達的存在下,將絲氨酸(中圖)或半胱氨酸(右圖)殘基突變成丙氨酸防止了硫醚-橋的酶形成,並使得肽對xa因子敏感。從三組獨立產生的細胞溶解產物,計算相對於未處理樣品(w/oxa)的xa因子抗性(信號保留[%])。
圖2.含有nisa-前導的前體肽與piiic-端的融合物是酶環化(形成硫醚-橋)的底物,並隨後展示在噬菌體顆粒上
(a)使用將具有aswiegrwcn-基序的含有nisa-前導序列的相同肽(seq-id.:1)與piii的c-端或n-端融合的噬菌粒,產生噬菌體顆粒,並進行xa因子-切割報導基因檢驗。在nisb/nisc共表達的存在下產生、並在噬菌體上展示的c-端融合物對xa因子-切割具有很大的抗性,表明了生產細胞中的酶修飾以及隨後併入到噬菌體顆粒中(左圖)。相反地,即使在nisb/nisc共表達的存在下產生,噬菌體上展示的相同肽的n-端融合物也對xa因子敏感(右圖)。(b)即使在nisb/nisc共表達的存在下,在c-端piii融合物中心肽中將絲氨酸(左圖)或半胱氨酸(右圖)殘基突變成丙氨酸,也防止了酶促硫醚-橋的形成,並使展示的肽對xa因子敏感。從三組獨立產生的噬菌體樣品,計算相對於未處理樣品(w/oxa)的xa因子抗性(信號保留[%])。(c)產生與麥芽糖結合蛋白(mbp)的c-端或n-端融合的表達含有(a)中所示的nisa-前導序列的相同前體肽的大腸桿菌菌株的細胞溶解產物,並對其進行xa因子-切割報導基因檢驗。當在nisb/nisc共表達的存在下產生時,前體肽與mbpc-端的融合物對xa因子-切割的抗性很大,證實了酶修飾。在不存在nisb/nisc共表達的情況下,檢測不到相同的肽融合物(n.d.),這可能說明有迅速的轉換(左圖)。無論有沒有nisb/nisc共表達,前體肽與mbp的n-端融合物累積至高的水平,但不能被酶修飾,如xa因子-切割檢驗所判斷(右圖)。結果表明,前體肽與載體蛋白c-端的融合是廣泛適用的,並與n-端融合相反,其通過酶機制支持有效的修飾。
圖3.含有proca-前導的前體肽與piiic-端的融合物是酶環化(形成硫醚-橋)的底物,並隨後在噬菌體顆粒上展示
使用將具有aswiegrwcn-基序的含有proca-前導的肽(seq-id.:1;s/c;或關聯物t/c和s/a衍生物)與piiic-端融合的噬菌粒產生噬菌體顆粒,並對其進行xa因子-切割報導基因檢驗。在procm酶共表達的存在下產生的含有s/c(左圖)或t/c(右圖)殘基的c-端肽融合物表現出對xa因子-切割的抗性,表明分別由脫氫丙氨酸(s/c)和脫氫氨基丁酸(t/c)酶促形成硫醚-橋,並且隨後併入到噬菌體顆粒中。相反地,具有s/a殘基的融合物不是形成硫醚的底物,即使在procm的存在下產生也保持對xa敏感(右圖)。從三組獨立產生的噬菌體樣品,計算相對於未處理樣品(w/oxa)的xa因子抗性(信號保留[%])。
圖4:腸桿菌(enterobacteria)噬菌體m13g3p(piii)和g8p(pviii)蛋白質的野生型胺基酸序列
提供piii(seq-id.:2;uniprot-id:p69168)和pviii(seq-id.:3;uniprot-id:p69541)蛋白質的序列,信號肽(黑體)和跨膜結構域序列(黑體,下劃線)突出顯示。
圖5:在piiic端上含有nisa-和proca-前導序列並且環大小可變的肽融合物的酶環化(形成硫醚-橋)和噬菌體展示
(a)使用將分別具有aswiegrecn-(seq-id.:11)、aswaaiegraecn-(seq-id.:12)、aswaaaiegraaaecn-(seq-id.:13)或aswaagaaiegraagaaecn-基序(seq-id.:14)的含有nisa-前導的肽(分別為構建物i,i+7、i,i+10、i,i+13和i,i+17;xa因子切割位點加下劃線)與piiic-端融合的噬菌粒,產生噬菌體顆粒,並進行xa因子-切割報導基因檢驗。不在nisb/c酶共表達的存在下產生的c-端肽融合物表現出對獨立於環大小的xa因子-切割的抗性,其由絲氨酸/半胱氨酸間隔控制。從三組獨立產生的噬菌體樣品,計算相對於未處理樣品(w/oxa)的xa因子抗性(信號保留[%])。
(b)與(a)相同,但使用含有proca-前導序列的piii-肽融合物,在存在或不存在procm酶共表達下產生。
具體實施方式
定義
術語「噬菌體」是指形成由含有噬菌體複製所需核酸的蛋白質外被組成的包裝的細菌病毒。核酸可以是dna或rna,雙鏈的或單鏈的,線性的或環狀的。例如噬菌體λ或絲狀噬菌體(例如ml3、fd或fl)的噬菌體是本領域普通技術人員公知的。術語「噬菌體顆粒」是指根據本發明的顆粒,即展示環肽的顆粒。在噬菌體組裝過程中,外被蛋白可以包裝不同的核酸序列,只要其包括包裝信號即可。
術語「核酸」是指單鏈或雙鏈形式的脫氧核糖核苷酸或核糖核苷酸及其聚合物。該術語包括含有已知的核苷酸類似物或修飾的骨架殘基或鍵的核酸,其是合成的、天然的和非天然的,具有與參考核酸類似的結合特性,並且以與參考核苷酸類似的方式被代謝。除非另外指出,特定的核酸序列還隱含地包括其保守修飾的變體(例如,簡併密碼子置換物)和互補序列,以及明確指出的序列。具體地,如下文所詳述,通過產生其中第三位置的一個或多個所選(或全部)密碼子被混合-鹼基和/或脫氧肌苷鹼基取代的序列,可以實現簡併密碼子置換(batzer等人(1991)nucleicacidres.19:5081;ohtsuka等人(1985)j.biol.chem.260:2605-2608;和rossolini等人(1994)mol.cell.probes8:91-98)。本文公開的特定核酸序列或載體具有在噬菌體或噬菌體顆粒的組裝過程中被噬菌體外被蛋白包裝的能力。優選地,所述核酸序列或載體源自噬菌體的天然基因組,例如,在絲狀噬菌體的情況下,包括噬菌體和噬菌粒載體。後者是除質粒特徵以外還含有包裝信號和噬菌體複製起源的質粒。
術語「肽」是指具有少於或等於50個胺基酸的分子。
術語「(多)肽」是指具有多於50個胺基酸的分子,其由一種或多種經由肽鍵連接的多個(即2個或更多個)胺基酸鏈組成。
術語「蛋白質」是指如下(多)肽:其中至少一部分(多)肽通過在(多)肽鏈內和/或其之間形成二級、三級或四級結構具有或能夠取得確定的三維排列。該定義包括多種蛋白質,例如天然的或至少部分人工的蛋白質,以及全蛋白質的片段或結構域,只要這些片段或結構域能夠取得上述的確定三維排列即可。
術語「硫醚」或「硫醚-橋」是指在各個分子中硫原子與兩個不同的碳原子或雜原子結合。在一實施方式中,硫醚-橋在一個或多個絲氨酸或蘇氨酸殘基的翻譯後脫水以及將所述脫水殘基與半胱氨酸偶聯之後形成。在一實施方式中,硫醚-橋是羊毛硫氨酸-或甲基羊毛硫氨酸-橋。羊毛硫氨酸是化學式為(hooc-ch(nh2)-ch2-s-ch2-ch(nh2)-cooh)的非蛋白原胺基酸,其由兩個在其β-碳原子上通過硫醚-橋交聯的丙氨酸殘基組成。甲基羊毛硫氨酸是化學式為hooc-ch(nh2)-ch(ch3)-s-ch2-ch(nh2)-cooh的非蛋白原胺基酸。
術語「賴丙氨酸-橋」是指脫氫丙氨酸與賴氨酸殘基之間的相互作用。在本文中,通過酶法或非酶法(例如,通過調節ph)引起賴丙氨酸-橋。「賴丙氨酸」是指修飾的胺基酸n6-(dl-2-氨基-2-羧基乙基l)-l-賴氨酸。
如本文所用,術語「分子內鍵」是指肽序列內胺基酸側鏈之間的共價鍵,不包括分子外(外源性)結構,並排除化學處理,例如形成二硫橋(例如通過還原反應)、環加成或staudinger反應。在此,分子內鍵可以通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯形成。在一實施方式中,所述一個或多個脫水殘基是脫氫丙氨酸(dha)或脫氫氨基丁酸(dhb)。在此,分子內鍵可以通過賴丙氨酸-橋或硫醚-橋形成。在本公開內容的一個實施方式中,分子內鍵通過羊毛硫氨酸-橋形成。在本公開內容的另一實施方式中,分子內鍵通過甲基羊毛硫氨酸-橋形成。在本公開內容的實施方式中,一個或多個分子內鍵在肽序列內形成。在本公開內容的實施方式中,肽序列內的分子內鍵形成穩定化環結構。在一實施方式中,所述一個或多個分子內鍵通過酶法形成。在一實施方式中,所述一個或多個分子內鍵通過羊毛硫肽合成酶形成。所述一個或多個分子內鍵通過環化酶形成。在一實施方式中,所述環化酶是lanc型環化酶或雙功能lanm型酶或多功能lankc或lanl型酶。在另一實施方式中,所述lanc型環化酶是nisc(uniprot編號:q03202)、spac、mibc、pepc、epic或其功能等同物。在另一實施方式中,所述雙功能lanm型酶是procm(編號np_894083)、lctm、mutm、bovm、lanm1/2、cinm、halml/2、cyanml-4或其功能等同物。在另一實施方式中,所述一個或多個分子內鍵通過非酶法形成。在另一實施方式中,所述一個或多個分子內鍵在鹼性條件下形成。在另一實施方式中,所述一個或多個分子內鍵在弱鹼性條件下進行。在另一實施方式中,所述一個或多個分子內鍵在鹼性條件下形成。在另一實施方式中,所述鹼性條件為ph7.5、ph8、ph8.5、ph9、ph9.5、ph10、ph10.5、ph11、ph11.5、ph12、ph12.5、ph13、ph13.5或ph14。在另一實施方式中,所述一個或多個分子內鍵在鹼性條件下通過酶法形成,例如通過羊毛硫肽合成酶。
在本文中,術語「環肽」是指具有通過一個或多個分子內鍵形成的二級結構的一段胺基酸、肽或多肽。並不需要整段胺基酸或肽或多肽是環狀的。在本公開內容的實施方式中,環肽是單環或多環的肽。在另一實施方式中,環肽包括例如天然或人工肽的肽,以及作為全蛋白質片段或結構域的肽。在另一實施方式中,環肽是醯胺化環肽。
術語「多環」或「多環結構」是指具有至少2、3、4或5個分子內鍵的結構。取決於所用肽的長度,根據本公開內容,可以實現更加複雜的二級肽結構。
術語「前體環肽」是指能夠形成根據本公開內容的環肽的一段胺基酸、肽或多肽。更具體地,根據本公開內容的前體環肽包括至少一個或多個絲氨酸或蘇氨酸和至少一個或多個半胱氨酸或賴氨酸,以形成分子內鍵。
術語「脫水殘基」是指經歷包括從反應分子上失去水分子的化學反應的修飾的胺基酸殘基。在一實施方式中,「脫水殘基」是脫水絲氨酸或脫水蘇氨酸。在另一實施方式中,「脫水殘基」是脫氫丙氨酸(dha)或脫氫氨基丁酸(dhb)。在一實施方式中,一個或多個絲氨酸或蘇氨酸的脫水通過羊毛硫肽合成酶進行。在一實施方式中,一個或多個絲氨酸或蘇氨酸的脫水通過脫水酶進行。在一實施方式中,所述酶脫水通過lanb型脫水酶或者通過雙功能lanm型酶或多功能lankc或lanl型酶進行。在一實施方式中,所述lanb型脫水酶是nisb(uniprot編號:p20103)、epib、spab、mibb、pepb或其功能等同物。在另一實施方式中,所述雙功能lanm型酶是procm(編號np_894083)、lctm、mutm、bovm、lanml/2、cinm、halml/2、cyanml-4或其功能等同物。
如本文所述,術語「前導」或「前導序列」是指翻譯後修飾(ptm)酶的識別基序。在本公開內容一實施方式中,前導序列由翻譯後修飾(ptm)酶識別。在本公開內容一實施方式中,前導序列是羊毛硫肽合成酶識別的序列。在另一實施方式中,前導序列具有可以源自由翻譯後修飾(ptm)酶識別的前導序列的共有基序。在本公開內容另一實施方式中,前導序列是源自lana前體肽的序列。在本公開內容另一實施方式中,前導序列具有可以源自lana前體肽的共有基序。在本公開內容另一實施方式中,前導序列是由lanb型脫水酶、lanc型環化酶和/或雙功能lanm型酶或多功能lankc或lanl型酶識別的序列。
如本文所用,術語「翻譯後修飾酶」或「ptm酶」是指引發翻譯的肽(特別是在生物活性肽的生物合成中作為處理機制一部分的修飾天然核糖體肽)的結構變化的酶。該類別包括多種類型的酶,包括羧酸-胺連接酶、環化酶、脫氫酶、環化脫水酶、脫羧酶、差向異構酶、羥化酶、肽酶、脫水酶、裂解酶、激酶、轉移酶、酯化酶、加氧酶和異構酶,特別是形成羊毛硫氨酸鍵的酶、形成細胞溶素的酶、形成cyanobactin的酶、形成硫肽的酶、形成芋螺肽(conopeptide)肽的酶、形成microviridin的酶、形成大環寡肽(cyclotide)的酶、形成細菌素的酶和形成subtilosin的酶。優選地,本文使用的ptm酶是羊毛硫肽合成酶。優選地,本文使用的ptm酶是脫水酶、環化酶或包括脫水酶或裂解酶/激酶和環化酶活性的雙-或多功能酶。更優選地,本文使用的ptm酶是lanb型脫水酶、lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶或其功能等同物。
術語肽或蛋白質的「功能等同物」是指共享該肽或蛋白質一種或多種、優選全部功能者。優選地,這些功能是生物功能,優選酶功能,例如脫水酶和/或環化酶活性。
術語「噬菌體顆粒表面」是指噬菌體顆粒與顆粒包含於其中並可接近的介質接觸的部分。該表面通過作為噬菌體外被部分的蛋白質(顆粒蛋白質外被的膜)確定,其在適當的宿主細胞中在噬菌體產生過程中組裝。
噬菌體展示描述了如下選擇技術:其中肽或蛋白質變體文庫在噬菌體病毒子的外部表達,而編碼各變體的遺傳材料位於內部。這在各變體蛋白質序列和編碼其的dna之間建立了物理連接,使得可以通過稱作淘選的體外選擇方法基於對指定目標分子(抗體、酶、細胞-表面受體等)的結合親合力進行迅速區分。淘選以其最簡單的方式進行:在覆有靶標的板(或珠)上孵育噬菌體-展示的肽文庫,洗去未結合的噬菌體,並洗脫特異性結合的噬菌體。然後將洗脫的噬菌體進行擴增,採取額外的結合/擴增循環,以富集有利於結合序列的庫。若干輪之後,通過dna測序或elisa對單個克隆進行表徵。
術語「噬菌粒」是指具有細菌複製起源例如coie1和一拷貝噬菌體基因間區域的質粒載體。噬菌粒可以基於任何已知的噬菌體,包括絲狀噬菌體。質粒通常會含有用於抗生素抗性的選擇標記物。克隆到這些載體中的dna片段可以作為質粒繁殖。當提供具有產生噬菌體顆粒所必需的所有基因的帶有這些載體的細胞時,質粒複製的方式變化成滾環複製,以產生一鏈質粒dna鏈的多個拷貝以及包裝噬菌體顆粒。噬菌粒可以形成感染性或非感染性噬菌體顆粒。該術語包括含有作為基因融合物的與異源性多肽基因連接的噬菌體外被蛋白基因或其片段的噬菌粒,使得異源性多肽在噬菌體顆粒的表面上得以展示(sambrook等人417)。
術語「噬菌體載體」是指含有異源性基因並能夠複製的噬菌體的雙鏈可複製形式。噬菌體載體具有允許噬菌體複製並形成噬菌體顆粒的噬菌體複製源。優選地,噬菌體是絲狀噬菌體,例如m13、fl、fd、pf3噬菌體或其衍生物,λ-型噬菌體,例如λ、21、phi80、phi81.82、424.434等或其衍生物,杆狀病毒或其衍生物,t4噬菌體或其衍生物,t7噬菌體病毒或其衍生物。由細胞製備dna意味著從宿主細胞的分培養物分離質粒dna。常用的dna製備方法為sambrook等人的125-133部分中所述的大規模和小規模質粒製備。製備dna之後,可以通過本領域公知的方法對其進行提純,例如sambrook等人的140部分中所述的。
術語「外被蛋白」是指存在於噬菌體顆粒表面上的蛋白質或其至少一部分。就功能性而言,外被蛋白是在宿主細胞中的噬菌體組裝過程中與噬菌體顆粒締合、並且保持與組裝的噬菌體締合直至其感染另一宿主細胞的任意蛋白。在絲狀噬菌體的情形中,所述野生型蛋白質是基因iii蛋白質(piii)、基因vi蛋白質(pvi)、基因vii蛋白質(pvii)、基因viii蛋白質(pviii)和基因ix蛋白質(pix)。外被蛋白可以是主要外被蛋白,或者可以是次要外被蛋白。「主要」外被蛋白是以10個蛋白質拷貝或更多地存在於噬菌體外被中的外被蛋白,例如,主要外被蛋白pviii。主要外被蛋白可以每噬菌體數十、數百甚至數千個拷貝存在。次要外被蛋白以每噬菌體少於10個蛋白質拷貝存在,例如,次要外被蛋白piii。
術語「野生型外被蛋白」是指形成天然存在的噬菌體的外被的外被蛋白。包括密切相關的絲狀噬菌體成員例如fl、fd和ml3的序列是本領域普通技術人員公知的(參見,例如,kay等人,1996)。在絲狀噬菌體的情形中,所述野生型蛋白質是,例如,基因iii蛋白質(piii)、基因vi蛋白質(pvi)、基因vii蛋白質(pvii)、基因viii蛋白質(pviii)和基因ix蛋白質(pix)。在一實施方式中,本公開內容涉及一種方法,其中所述外被蛋白是噬菌體的野生型外被蛋白。
在進一步優選的實施方式中,所述外被蛋白是噬菌體野生型外被蛋白的截短變體,其中所述截短變體包括所述野生型外被蛋白的至少一部分,所述部分引起所述外被蛋白結合到噬菌體顆粒蛋白質外被中的至少該部分。
術語「截短變體」是指源自上文所稱的野生型蛋白質的蛋白質,其通過刪除至少一部分野生型序列而修飾。這包括多種變體,例如截短的基因iii(piii)或基因viii(pviii)蛋白質變體,其已經發現於噬菌體突變體中(crissman&smith,1984),或者已經在標準噬菌體展示方法中產生(例如,bass等人,1990;krebber,1996)。例如,所述截短的變體可以包括基因iii蛋白質(piii)或基因viii蛋白質(pviii)的ct結構域或者由其組成。為了識別根據本發明的截短變體,可以將檢測標記與變體融合,並設立檢驗,以確定變體是否結合到在變體的存在下形成的噬菌體顆粒的噬菌體外被中。
在再進一步優選的實施方式中,所述外被蛋白是噬菌體野生型外被蛋白的修飾變體,其中所述修飾變體能夠結合到噬菌體顆粒的蛋白質外被中。
用於實現根據本公開內容的野生型蛋白質的修飾的方法是本領域普通技術人員公知的,其包括標準克隆和/或誘變技術。用於構建編碼根據本公開內容的方法中使用的野生型蛋白質修飾變體的核酸分子的方法,用於構建包括所述核酸分子的載體、包括構建噬菌體和/或噬菌粒載體的方法,用於將所述載體引入到適當選擇的宿主細胞中,用於引起或允許所述修飾蛋白質表達的方法,均是本領域公知的(參見,例如,sambrook等人,1989;ausubel等人,1999;kay等人,1996)。為了識別根據本公開內容的修飾變體,可以將檢測標記與變體融合,並設立檢驗,以確定變體是否結合到在變體的存在下形成的噬菌體顆粒的噬菌體外被中。
在再進一步優選的方法中,所述噬菌體是絲狀噬菌體。絲狀噬菌體例如m13、fd或f1是本領域普通技術人員公知的。
在絲狀噬菌體的情形中,特別優選的方法是其中噬菌體的所述外被蛋白是野生型外被蛋白piii或源自於此。
進一步優選的方法其中噬菌體顆粒的所述外被蛋白是野生型外被蛋白piii或源自於此。優選地,與對應的野生型序列相比較,對應於野生型蛋白質的修飾蛋白質的這些部分表現出的胺基酸同一性超過大約40%,優選大約50%,優選大約60%,優選大約70%,優選大約80%,最優選大約90%。
術語指定多肽序列的「n-端」是始於指定多肽序列n-端殘基處或其附近的連續長度的指定多肽序列。指定多肽的n-端可以通過長度定義。類似地,術語指定多肽序列的「c-端」是始於指定多肽序列c-端殘基處或其附近的連續長度的指定多肽序列。指定多肽的c-端可以通過長度定義。在本公開內容一實施方式中,是指外被蛋白的c-端。在優選的實施方式中,外被蛋白的c端是位於所述外被蛋白跨膜結構域c-端的胺基酸或胺基酸序列。在另一實施方式中,外被蛋白的c-端是位於所述外被蛋白跨膜結構域c-端的胺基酸或胺基酸序列,其中所述外被蛋白是基因iii蛋白質(piii,seq-id.:2;uniprot:p69168)或基因viii蛋白質(pviii;seq-id.:3;uniprot:p69541)。在一實施方式中,基因iii蛋白質(piii)的c-端是胺基酸序列lrnkes(seq-id.:4)或其衍生物或修飾變體。在另一實施方式中,基因iii蛋白質(piii)的c-端位於跨膜結構域的c-端,並包括胺基酸序列lrnkes(seq-id.:4)的一個或多個胺基酸。在另一實施方式中,基因viii蛋白質(pviii)的c-端是胺基酸序列tskas(seq-id.:5)或其衍生物或修飾變體。在另一實施方式中,基因viii蛋白質(pviii)的c-端位於跨膜結構域的c-端,並包括胺基酸序列tskas(seq-id.:5)的一個或多個胺基酸。圖4顯示了基因iii蛋白質(piii)和基因viii蛋白質(pviii)的示意圖。
在本公開內容一實施方式中,蛋白質外被部件的c-端與環肽連接,並在噬菌體顆粒的表面上展示所述環肽。
在優選的實施方式中,展示(多)肽/蛋白質的噬菌體顆粒含有編碼(多)肽/蛋白質的核酸序列。
在本公開內容上下文中,術語「造成或允許表達」描述的是在使得核酸序列得以表達的條件下培養宿主細胞。用於構建編碼根據本公開內容的(多)肽/蛋白質的核酸分子的方法,用於構建包括所述核酸分子的載體的方法,用於將所述載體引入到適當選擇的宿主細胞中,用於造成或允許(多)肽/蛋白質表達的方法是本領域公知的(參見,例如,sambrook等人,1989;ausubel等人1999)。進一步公知的有用於在適當宿主細胞中引入產生子代噬菌體或噬菌體顆粒所需的遺傳材料的方法,用於造成或者允許所述子代噬菌體或噬菌體顆粒產生的方法(參見,例如,kay等人,1996)。造成或允許噬菌體顆粒產生的步驟可以要求使用適當的輔助噬菌體,例如,在用噬菌粒工作的情形中。
在另一實施方式中,本公開內容涉及包括根據本公開內容的核酸序列的載體。
在另一實施方式中,本公開內容涉及含有根據本公開內容的核酸序列或根據本公開內容的載體的宿主細胞。
在本公開內容上下文中,術語「宿主細胞」可以是若干在異源性蛋白質的產生中常用的任意一種,包括,但不限於,細菌,例如大腸桿菌(escherichiacoli)(ge等人,1995)或枯草桿菌(bacillussubtilis)(wu等人,1993),真菌,例如酵母(horwitz等人,1988;ridderetal.,1995)或絲狀真菌(nyyssonen等人,1993),植物細胞(hiatt&ma,1993;whitelam等人,1994),昆蟲細胞(potter等人,1993;ward等人,1995),或哺乳動物細胞(trill等人,1995)。
在再進一步優選的實施方式中,本公開內容涉及根據本公開內容的核酸序列編碼的、根據本公開內容的載體編碼的、或者根據本公開內容的宿主細胞產生的野生型噬菌體外被蛋白的修飾變體。修飾變體可以進一步包括用於克隆、用於表達或蛋白質轉運所需的胺基酸殘基。克隆所需的蛋白質殘基可以包括由具有限制性內切酶用識別序列的核酸序列編碼的殘基,引入其是為了能夠將核酸序列克隆到合適的載體中。表達所需的胺基酸序列可以包括導致(多)肽/蛋白質的溶解性或穩定性增加的殘基。蛋白質轉運所需的胺基酸殘基可以包括負責將修飾變體轉運至大腸桿菌周質的信號傳導序列,和/或有利於所述信號傳導序列的有效切割的胺基酸殘基。其他以上所稱的用於克隆、表達、蛋白質轉運、提純和/或檢測目的的胺基酸殘基為眾多本領域普通技術人員公知的部分。
「噬菌體顆粒的多樣收集物」也可以稱作「文庫」或「多元化」。在本公開內容上下文中,該文庫中的每個成員展示文庫的不同成員。在本公開內容上下文中,術語「多樣收集物」是指至少兩種在其組成、特性和/或序列方面至少部分不同的顆粒或分子的收集物。例如,環肽的多樣收集物是一組在其序列上至少一個胺基酸位置上不同的環肽。這樣的多樣收集物可以許多種方式得到,例如,通過編碼起始(多)肽/蛋白質的核酸序列的至少一個密碼子的隨機誘變,通過使用易錯pcr,以擴增編碼起始(多)肽/蛋白質的核酸序列,或者通過在根據本公開內容的方法中使用突變體菌株作為宿主細胞。這些以及另外的或可選的產生肽多樣收集物的方法是本領域普通技術人員公知的。
在本公開內容上下文中,術語「所需特性」是指多樣收集物中一成員應當具有並形成篩選和/或選擇多樣收集物基礎的預定特性。這些特性包括如下特性,例如,與靶標結合,阻斷靶標,激活靶標介導的反應,酶活性,以及本領域普通技術人員已知的其他特性。取決於所需特性的類型,本領域普通技術人員將能夠識別出進行篩選和/或選擇的形式和必要步驟。
最優選的方法是其中所述所需特性是與所關注的靶標結合。
所述的所關注靶標可以通過本領域普通技術人員公知的許多種方式呈現於噬菌體顆粒上展示的環肽的所述多樣收集物,例如,塗覆在用於固相生物淘選的表面上,與例如磁珠的顆粒連接用於在溶液中淘選,或者展示在細胞表面上用於全細胞生物淘選或在組織切片上進行生物淘選。已經與所述靶標結合的噬菌體顆粒可以通過許多種本領域普通技術人員公知的方法回收,例如,通過用適當的緩衝液洗脫,通過ph-或鹽梯度,或者通過使用可溶性靶標的特異性洗脫。
術語「在附近」是指從所述(多)肽/蛋白質的n-或c-端開始計數至多15個或更優選至多10個胺基酸的一段。
詳述和實施方式
本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接,並且其中所述環肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。
在一實施方式中,本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供帶有包括編碼前體環肽的核酸序列的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽的表達;
(c)使前體環肽內的一個或多個胺基酸殘基酶脫水;
(d)通過使一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯,形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
在一實施方式中,本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供帶有編碼前體環肽的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽的表達;
(c)使前體環肽內的一個或多個胺基酸殘基酶脫水;
(d)通過使一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯,引起或允許形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
在本公開內容一實施方式中,所述核酸序列進一步編碼噬菌體顆粒的外被蛋白和由翻譯後修飾(ptm)酶識別的前導序列。
在本公開內容一實施方式中,宿主細胞進一步具有一個或多個編碼翻譯後修飾(ptm)酶的核酸序列。在本公開內容另一實施方式中,宿主細胞進一步具有一個或多個編碼翻譯後修飾(ptm)酶的核酸序列,其中所述一個或多個核酸序列被人工引入到細胞中。
在一實施方式中,本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供具有編碼前體環肽的核酸序列、並具有一個或多個編碼翻譯後修飾(ptm)酶的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽和所述一種或多種翻譯後修飾(ptm)酶的表達;
(c)使前體環肽內的一個或多個胺基酸殘基酶脫水;
(d)通過使一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯,形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
在一實施方式中,本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供具有編碼前體環肽的核酸序列的具有一個或多個編碼翻譯後修飾(ptm)酶的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽和所述一種或多種翻譯後修飾(ptm)酶的表達;
(c)引起或允許前體環肽內的一個或多個胺基酸殘基的酶脫水;
(d)通過使一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯,引起或允許形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
在一實施方式中,步驟c)中的酶脫水由翻譯後修飾(ptm)酶引起。在一實施方式中,步驟d)中一個或多個分子內鍵的形成由翻譯後修飾(ptm)酶引起。在一實施方式中,步驟c)中的酶脫水和步驟d)中一個或多個分子內鍵的形成由翻譯後修飾(ptm)酶引起。在一實施方式中,步驟c)中的酶脫水和步驟d)中一個或多個分子內鍵的形成由一種翻譯後修飾(ptm)酶引起。在另一實施方式中,步驟c)中的酶脫水和步驟d)中一個或多個分子內鍵的形成通過不同的翻譯後修飾(ptm)酶引起。在另一實施方式中,步驟c)中的酶脫水由翻譯後修飾(ptm)酶引起,在步驟d)中一個或多個分子內鍵在鹼性條件下形成。另再一實施方式中,步驟c)中的酶脫水由翻譯後修飾(ptm)酶引起,在宿主細胞中產生噬菌體顆粒之後,一個或多個分子內鍵在鹼性條件下形成。
在本公開內容一實施方式中,所述翻譯後修飾(ptm)酶是羊毛硫肽合成酶。在本公開內容另一實施方式中,所述翻譯後修飾(ptm)酶是脫水酶、環化酶或包括脫水酶或裂解酶/激酶和環化酶活性的雙功能或多功能酶。在優選的實施方式中,本文使用的ptm酶是lanb型脫水酶、lanc型環化酶和/或雙功能lanm型酶或多功能lankc或lanl型酶或其功能等同物。在本公開內容另一實施方式中,所述翻譯後修飾(ptm)酶是脫水酶或裂解酶/激酶和/或環化酶。
在本公開內容一實施方式中,所述一種或多種脫水殘基是脫氫丙氨酸(dha)或脫氫氨基丁酸(dhb)。
在本公開內容一實施方式中,一個或多個分子內鍵通過羊毛硫肽合成酶或者在弱鹼性條件下形成。在本公開內容另一實施方式中,一個或多個分子內鍵通過lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶或者在弱鹼性條件下形成。在本公開內容一實施方式中,胺基酸殘基的脫水和分子內鍵的形成通過羊毛硫氨酸-或甲基羊毛硫氨酸-橋形成酶介導。
在本公開內容一實施方式中,一種或多種分子內鍵在肽或多肽序列內形成。在本公開內容一實施方式中,肽或多肽序列內的分子內鍵形成穩定化環結構。在本公開內容一實施方式中,一個或多個分子內鍵在翻譯後修飾的肽或多肽內形成。在本公開內容一實施方式中,一個或多個分子內鍵在包括一個或多個脫水殘基的肽或多肽內形成。在另一實施方式中,一個或多個分子內鍵在通過羊毛硫氨酸合成酶修飾的肽或多肽內形成。在另一實施方式中,一個或多個分子內鍵在通過脫水酶、環化酶或包括脫水酶或裂解酶/激酶和環化活性的雙功能或多功能酶修飾的肽或多肽內形成。在另一實施方式中,一個或多個分子內鍵在通過lanb型脫水酶、lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶修飾的肽或多肽內形成。在另一實施方式中,lanb型脫水酶是nisb(uniprot編號:p20103)、epib、spab、mibb、pepb或其功能等同物。在另一實施方式中,所述雙功能lanm型酶是來自原綠球藻(prochlorococcus)mit9313的procm(編號np_894083)或其密切相關的來自原綠球藻mit9303的procm(編號yp_001018107)的類似物、cyanml-4(編號yp_002485891、yp_002483601、yp_002484655、yp_002483742;來自藍絲菌屬(cyanothecesp.)pcc7425)、lctm、mutm、bovm、lanml/2、cinm、halml/2或其功能等同物。
在一實施方式中,所述一個或多個分子內鍵通過羊毛硫氨酸-或甲基羊毛硫氨酸-橋形成酶形成。在一實施方式中,所述一個或多個分子內鍵通過環化酶形成。在一實施方式中,所述環化酶是lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶。
在另一實施方式中,lanc型環化酶是nisc(uniprot編號:q03202)、spac、mibc、pepc、epic或其功能等同物。在另一實施方式中,所述雙功能lanm型酶是來自原綠球藻mit9313的procm(編號np_894083)或其密切相關的來自原綠球藻mit9303的procm(編號yp_001018107)的類似物、cyanml-4(編號yp_002485891、yp_002483601、yp_002484655、yp_002483742;來自藍絲菌屬pcc7425)、lctm、mutm、bovm、lanml/2、cinm、halml/2或其功能等同物。
在另一實施方式中,所述一個或多個分子內鍵在弱鹼性條件下形成。在另一實施方式中,所述弱鹼性條件是ph7.5-11、ph8-11、ph9-11、ph10-11、ph7.5-9、ph8-9或ph9-10。在另一實施方式中,所述弱鹼性條件為ph7.5、ph8、ph8.5、ph9、ph9.5、ph10、ph10.5、或ph11或其各自之間的範圍。
在本公開內容另一實施方式中,一個或多個分子內鍵是硫醚-橋。在本公開內容另一實施方式中,一個或多個分子內鍵是羊毛硫氨酸-橋或甲基羊毛硫氨酸-橋。在本公開內容另一實施方式中,一個或多個分子內鍵是賴丙氨酸-橋。
在本公開內容一實施方式中,環肽是翻譯後修飾的肽或多肽。在另一實施方式中,環肽是含有硫醚-橋的肽或多肽。在另一實施方式中,環肽是含有羊毛硫氨酸-橋的肽或多肽、含有甲基羊毛硫氨酸-橋的肽或多肽或者含有賴丙氨酸-橋的肽或多肽。在本公開內容另一實施方式中,環肽是單環的或多環的。
在一實施方式中,本公開內容涉及一種在噬菌體顆粒的表面上展示環肽的方法,其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接,並且其中所述環肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵,該方法包括以下步驟:
(a)提供帶有包括編碼前體環肽的核酸序列的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽的表達;
(c)形成一個或多個分子內鍵;和
(d)在所述宿主細胞中產生噬菌體顆粒。
在本公開內容一實施方式中,所述外被蛋白是噬菌體的野生型外被蛋白。在另一實施方式中,噬菌體顆粒的所述外被蛋白是或者源自於野生型外被蛋白piii或野生型外被蛋白pviii。
在本公開內容一實施方式中,所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。在另一實施方式中,所述環肽和所述噬菌體外被蛋白的c-端物理締合。在另一實施方式中,所述環肽經由基因融合或者經由通過一個或多個人工引入的半胱氨酸形成的二硫鍵與所述噬菌體顆粒外被蛋白的c-端連接。
在本公開內容一實施方式中,所述環肽經由基因融合或者經由通過一個或多個人工引入的半胱氨酸形成的二硫鍵與所述噬菌體顆粒外被蛋白的c-端連接。在另一實施方式中,所述環肽經由基因融合與所述噬菌體顆粒外被蛋白的c-端連接。在本公開內容另一實施方式中,所述環肽經由通過一個或多個人工引入的半胱氨酸形成的二硫鍵與所述噬菌體顆粒外被蛋白的c-端連接。
在本公開內容一實施方式中,所述噬菌體是絲狀噬菌體。
在本公開內容一實施方式中,所述環肽包括至多500、至多400、至多300、至多200、至多100、至多90、至多80、至多70、至多60、至多50、至多40、至多30、至多20或至多10個胺基酸。
在本公開內容另一實施方式中,所述環肽是醯胺化環肽。在另一實施方式中,所述醯胺化環肽在所述環肽的c-端包括醯胺部分。在本公開內容另一實施方式中,所述環肽通過c-端醯胺化進行翻譯後修飾。在另一實施方式中,要醯胺化修飾的胺基酸在提供醯胺基的甘氨酸之後。醯胺化包括,例如,第一反應步驟,其中將甘氨酸氧化形成α-羥基-甘氨酸。氧化的甘氨酸切割成c-端醯胺化的肽和n-乙醛酸化肽。c-端醯胺化對於許多肽例如神經肽和激素來說可能是至關重要的。
在一實施方式中,本公開內容涉及一種能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸編碼:
(a)所述噬菌體顆粒的外被蛋白,
(b)由翻譯後修飾(ptm)酶識別的前導序列,和
(c)前體環肽,
其中前體環肽位於所述噬菌體顆粒外被蛋白的c-端處,並且
其中所述前體環肽能夠通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成分子內鍵。
在本公開內容一實施方式中,能夠在噬菌體顆粒的表面上展示環肽的所述核酸序列還編碼信號序列。
在一實施方式中,本公開內容涉及一種載體,其包括能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸編碼:
(a)所述噬菌體顆粒的外被蛋白,
(b)由翻譯後修飾(ptm)酶識別的前導序列,和
(c)前體環肽,
其中前體環肽位於所述噬菌體顆粒外被蛋白的c-端處,並且
其中所述前體環肽能夠通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成分子內鍵。
在一實施方式中,本公開內容涉及一種能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸序列從n-端到c-端具有以下排列:
n-(噬菌體外被蛋白)-(由翻譯後修飾(ptm)酶識別的前導序列)-(前體環肽)-c。
在一實施方式中,本公開內容涉及一種載體,其包括能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸從n-端到c-端具有以下排列:
n-(噬菌體外被蛋白)-(由翻譯後修飾(ptm)酶識別的前導序列)-(前體環肽)-c,其中n是n-端,c是c-端。
在一實施方式中,本公開內容涉及一種能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸從n-端到c-端具有以下排列:
n-(信號序列)-(噬菌體外被蛋白)-(由翻譯後修飾(ptm)酶識別的前導序列)-(前體環肽)-c。
在一實施方式中,本公開內容涉及一種載體,其包括能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸從n-端到c-端具有以下排列:
n-(信號序列)-(噬菌體外被蛋白)-(由翻譯後修飾(ptm)酶識別的前導序列)-(前體環肽)-c,其中n是n-端,c是c-端。
在本公開內容另一實施方式中,載體還包括一個或多個編碼輸出信號的核酸序列。在本公開內容另一實施方式中,載體還包括一個或多個編碼誘導型啟動子的核酸序列。在一實施方式中,本公開內容涉及一種包括本文所公開的核酸序列或載體的宿主細胞。
在另一實施方式中,本公開內容涉及一種載體,其包括能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸編碼噬菌體外被蛋白,其中所述噬菌體外被蛋白在由翻譯後修飾(ptm)酶識別的前導序列附近編碼。
在另一實施方式中,本公開內容涉及一種載體,其包括能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中由翻譯後修飾(ptm)酶識別的前導序列在前體環肽附近編碼。
在另一實施方式中,本公開內容涉及一種包括核酸序列的載體,其中在所述核酸序列上,噬菌體外被蛋白在由翻譯後修飾(ptm)酶識別的前導序列附近編碼,並且所述前導序列在前體環肽附近編碼。在本公開內容一實施方式中,在附近是指由對應的核酸三聯體編碼的15、14、13、12、11、10、9、8、7、6、5、4、3、2或1個胺基酸。
在另一實施方式中,所述前體環肽包括至少一個或多個絲氨酸或蘇氨酸和一個或多個半胱氨酸或賴氨酸,以形成一個或多個分子內鍵。在另一實施方式中,噬菌體顆粒的所述外被蛋白被作為野生型外被蛋白piii或野生型外被蛋白pviii的或由其衍生的核酸編碼。
在一實施方式中,本公開內容涉及通過本文所公開的方法得到的在其表面上展示環肽的噬菌體顆粒。在一實施方式中,本公開內容涉及在其表面上展示環肽的噬菌體顆粒,其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接,並且其中所述環肽包括分子內鍵。在一實施方式中,本公開內容涉及在其表面上展示環肽的噬菌體顆粒,其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接,並且其中所述環肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。
在另一實施方式中,所述噬菌體顆粒還包括載體,該載體包括一個或多個編碼能夠形成所述環肽的前體環肽的核酸序列。在另一實施方式中,所述噬菌體顆粒包括本文所公開的載體。
在一實施方式中,本公開內容涉及本文所公開的噬菌體顆粒的多樣收集物。在一實施方式中,本公開內容涉及噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽,並且其中所述多肽包括分子內鍵。在另一實施方式中,所述噬菌體顆粒中的每一個展示環肽多樣收集物中的環肽,其中所述環肽包括分子內鍵。在一實施方式中,本公開內容涉及噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。在另一實施方式中,所述噬菌體顆粒中的每一個展示環肽多樣收集物中的環肽,其中所述環肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。
在一實施方式中,本公開內容涉及一種獲得具有所需特性的環肽的方法,其包括:
(a)提供本文公開的噬菌體顆粒的多樣收集物;和
(b)篩選所述多樣收集物和/或從所述多樣收集物中選擇,以得到至少一種展示具有所述所需特性的環肽的噬菌體顆粒。
在一實施方式中,本公開內容涉及一種獲得具有所需特性的環肽的方法,其包括:
(a)提供噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵;和
(b)篩選所述多樣收集物和/或從所述多樣收集物中選擇,以得到至少一種展示具有所述所需特性的環肽的噬菌體顆粒。
在本公開內容一實施方式中,所述的所需特性是與所關注的靶標結合。
在另一實施方式中,本公開內容涉及一種獲得具有所需特性的環肽的方法,其包括:
(a)提供噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵;和
(b)篩選所述多樣收集物和/或從所述多樣收集物中選擇,以得到至少一種展示具有所述所需特性的環肽的噬菌體顆粒,其中步驟(b)還包括:
(ba)使噬菌體顆粒的所述多樣收集物與所關注的靶標接觸;
(bb)洗脫沒有與所關注的靶標結合的噬菌體顆粒;和
(bc)洗脫與所關注的靶標結合的噬菌體顆粒。
在一實施方式中,本公開內容涉及一種獲得具有所需特性的環肽的方法,其包括:
(a)提供噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽多樣收集物中的環肽,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵;和
(b)篩選所述多樣收集物和/或從所述多樣收集物中選擇,以得到至少一種展示具有所述所需特性的環肽的噬菌體顆粒。
在本公開內容一實施方式中,所述的所需特性是與所關注的靶標結合。
在另一實施方式中,本公開內容涉及一種獲得具有所需特性的環肽的方法,其包括:
(a)提供噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒展示與所述噬菌體顆粒外被蛋白的c-端連接的環肽多樣收集物中的環肽,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵;和
(b)篩選所述多樣收集物和/或從所述多樣收集物中選擇,以得到至少一種展示具有所述所需特性的環肽的噬菌體顆粒,其中步驟(b)還包括:
(ba)使噬菌體顆粒的所述多樣收集物與所關注的靶標接觸;
(bb)洗脫沒有與所關注的靶標結合的噬菌體顆粒;和
(bc)洗脫與所關注的靶標結合的噬菌體顆粒。
lan-酶
目前,若干羊毛硫氨酸-或甲基羊毛硫氨酸-橋形成酶及其基因是已知的:
1.lanb型脫水酶顯示出構成設計為催化絲氨酸和蘇氨酸脫水步驟的酶的c-端。
2.lanc型環化酶催化半胱氨酸硫醇的加成。環化酶組分lanc是鋅金屬蛋白,其結合的金屬設計為激活親和加成的硫醇底物。
3.lanm型酶是雙功能的脫水酶和環化酶。它在羊毛硫抗生素合成的過程中負責前體肽的脫水和環化。
4.land氧化性脫羧酶型酶催化從表皮纖維的c-端內消旋羊毛硫氨酸的半胱氨酸殘基上除去兩個還原當量,以形成-c=c-雙鍵。
5.lankc型多功能酶含有裂解酶、激酶和環化酶結構域,但在環化酶結構域中缺少標記鋅-配體。
6.lanl型多功能酶含有裂解酶、激酶和含有金屬配體的環化酶結構域。
7.lanp型肽酶從羊毛硫抗生素上切割前導肽。
8.lant型肽酶,與abc轉運體融合;前體肽的切割由作為分泌過程一部分的轉運體介導。
9.ltnm和ltnj型脫水酶和脫氫酶涉及d-丙氨酸的形成。
10.cinx在肉桂黴素的生物合成過程中使天冬醯胺羥基化。
例如,在羊毛硫抗生素(包括羊毛硫氨酸-橋或甲基羊毛硫氨酸-橋的抗微生物肽)的形成中,經描述羊毛硫抗生素-合成酶組裝在膜結合複合物中(siegers等人1996.j.biol.chem.271,12294-12301;kiesau等人1997.j.bacteriol.179,1475-1481;sahl等人1998.annu.rev.microbiol.52:41-7)。該複合物由羊毛硫抗生素轉運體(lant)、脫水酶(lanb;也稱作脫水酶)和環化酶(lanc)組成。在一些羊毛硫抗生素的情形中,雙功能酶(lanm)進行脫水和環化兩個步驟。在核糖體合成的前體肽中n-端羊毛硫抗生素前導肽是用於羊毛硫抗生素酶的識別信號,其始於脫水酶或既進行脫水也進行成環的酶。前導肽被認為與羊毛硫抗生素複合物結合,以使前體肽密切接近於羊毛硫抗生素酶。現有文獻(參見,例如,wo2006/062398)公開了若干羊毛硫抗生素前導肽及其用途,例如,用於融合蛋白中,以產生要通過脫水酶進行翻譯後脫水的所關注的肽。根據wo2006/062398,要修飾的前導肽或肽前面是非羊毛硫抗生素輸出信號,如sec輸出信號。輸出信號或前導肽可以通過細胞錨定序列(例如lptx-分選酶識別基序)隔開。
本領域技術人員可以使用基因組挖掘策略識別新的前導肽及其相關的修飾lan-酶,如最近所詳述(qizhang,xiaoyang,huanwang,wilfreda.vanderdonk(2014)acschem.biol.,2014,9,2686-2694)。lan-酶的特徵在於序列相似性、類似的結構域結構和高度保守的基序。例如,迄今已知若干lan-酶在環化酶結構域含有「chg」基序,其支持活性位點鋅離子的結合。識別出其他者含有「ccg」基序,其支持zn2+配位。使用lan-酶序列和保守基序作為查詢,可以通過blastp搜索在其他有機體中識別出新的lan-酶。一旦識別出新的lan-酶,其相關的底物lana前體肽很容易檢測,因為它們通常在相同基因簇中在附近編碼,並且特徵在於短的開放閱讀框和與其他lana的序列相似性。
前導序列
可以使用任意類型的前導序列用於實施本公開內容,只要其可以被脫水酶識別即可。在另一實施方式中,該前導序列也可以被可以形成羊毛硫氨酸-或甲基羊毛硫氨酸-橋的環化酶識別。前導序列的胺基酸序列可獲自公開資料庫和出版物,包括以下文獻中所述的前導序列和前導共有序列:plata.等人2013currproteinpeptsci.2013mar;14(2):85-96和plata.等人;applenvironmicrobiol.2011jan;77(2):604-11。
在一實施方式中,所述前導序列是由lanb型脫水酶、lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶識別的前導序列或者具有其共有基序。
在另一實施方式中,本公開內容的前導序列具有來自lana前體肽的前導序列或其共有基序,其可以自衍生自源自lana前體肽的已知前導序列的胺基酸序列對比。在另一實施方式中,所述前導序列是來自lana前體肽的前導序列,或者是衍生自lana前體肽的前導序列,或者是帶有來自lana前體肽的共有基序的前導序列,並且其中所述前導序列由羊毛硫肽合成酶識別。在另一實施方式中,所述前導序列是來自lana前體肽的前導序列,或者是衍生自lana前體肽的前導序列,或者是帶有來自lana前體肽的共有基序的前導序列,並且其中所述前導序列由lanb型脫水酶、lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶識別。
lana前體肽的胺基酸序列可獲自公開資料庫,其包括以下lana前體肽:
nisa(乳酸鏈球菌素,乳酸乳球菌)
proca(prochlorosin,原綠球藻mit9313;原綠球藻mit9303)
spas(枯草菌素;枯草芽孢桿菌(bacillussubtilis)atcc6633)
lcta(乳鏈球菌素(lacticin)481;乳酸乳球菌乳酸亞種)
muta(變鏈素(mutacin)ii,鏈球菌突變體)
miba(microbisporicin,microbisporacorallina)
bova(羊毛硫細菌素(bovicin)hj50;牛鏈球菌(streptococcusbovis)hj50)
lana1/2(lichenicidin,地衣芽孢桿菌(bacilluslicheniformis))
cina(肉桂黴素,streptomycescinnamoneuscinnamoneusdsm40005)
hala1/2(haloduracin,嗜鹼芽孢桿菌(bacillushalodurans))
cyana(未命名,藍絲菌屬pcc7425)
pep5(表皮葡萄球菌(staphylococcusepidermidis))
epia(表皮纖維,表皮葡萄球菌)
wo2006/062398的表1a和1b顯示了這些前導肽的示例性對比。本領域技術人員將能夠從對比的序列得到共有基序,例如,使用公開或市售的對比軟體,例如alignx或vectornti。優選地,前導序列共有基序源自至少5個、更優選至少10個、最優選至少15個已知前導肽序列的對比。由此得到的共有基序隨後可以使用本領域已知的方法進行前導肽活性的驗證,即被脫水酶和絲氨酸或蘇氨酸脫水用本領域已知的方法進行識別。指定目標序列的脫水可以使用maldi-tofms監測。
前導肽共有序列可以包括多種共有基序,例如,共有基序x1-d/e-e-v/l-s/t-d/e-x2-e-l-d/e,其中x1是任意疏水胺基酸,其中x2是任意胺基酸。例如,其包括序列leevseqeld(seq-id.:7)。在另一實施方式中,前導序列包括共有基序f-d/e/n-l-d/e/n-x3,其中x3是l、i或v。例如,其包括序列lfdldl(seq-id.:8)或fnldv(seq-id.:9)。例如,前導序列還可以含有共有i/l-l/f-d/e/n-l-q-d/n/a/s/t-l/m-d/e,其包括ilelqnld(seq-id.:10)。前導序列可以由共有序列例如fnldv(seq-id.:9)、之後在共有序列和要修飾的前體肽之間的間隔序列組成。該間隔序列使得要修飾的部分處於各個酶的催化中心所及範圍以內(annechienplat,leond.kluskens,annekekuipers,rickrink,gertn.moll(2010)),n-端結構域和間隔區對於乳酸鏈球菌素前導肽的功能性是充分的(then-terminaldomainandaspaceraresufficientforfunctionalityofthenisinleaderpeptide).appl.environ.microbiol.77,604-611)。
基於序列相似性將許多ii類lana前導肽歸為n11p、tigr03898和n11p族,而其他目前缺乏族分類。在另一實施方式中,前導序列源自於proca前導,其目前包括29個在原綠球藻mit9313中編碼的高度保守的成員和另外15個在原綠球藻mit9303中編碼的成員,其均被歸為前導肽的n11p族並且是在關聯菌株中編碼的單修飾procm酶的底物。大多數proca前導序列顯著地長(超過60個胺基酸),並且由於總體的高保守性,不具有清晰的最小共有基序。但是,將proca2.8的親本63個殘基前導序列n-端截短成23個殘基的版本充分支持procm-介導的前體肽的酶修飾,這說明前導序列的大部分是非必要的,並且可以很容易地得到功能性最小序列。
在另一實施方式中,前導序列是nisa(乳酸鏈球菌肽,乳酸乳球菌)或proca(prochlorosin,原綠球藻mit9313或原綠球藻mit9303)前導序列,或者具有可以源自於nisa或proca前導序列的共有基序。
本文公開的其他實施方式
本文公開的其他實施方式:
1.一種在噬菌體顆粒的表面上展示環肽的方法,其包括以下步驟:
(a)提供帶有編碼前體環肽的核酸序列的宿主細胞;
(b)引起或允許所述前體環肽的表達;
(c)使前體環肽內的一個或多個胺基酸殘基酶脫水;
(d)通過使一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯,形成一個或多個分子內鍵,從而形成環肽;和
(e)在所述宿主細胞中產生噬菌體顆粒,其中所述噬菌體顆粒在表面上展示所述環肽,並且其中所述環肽與所述噬菌體顆粒外被蛋白的c-端連接。
2.根據實施方式1所述的方法,其中所述核酸序列進一步編碼噬菌體顆粒的外被蛋白和由翻譯後修飾(ptm)酶識別的前導序列。
3.根據前述實施方式之一所述的方法,其中宿主細胞進一步具有一個或多個編碼翻譯後修飾(ptm)酶的核酸序列。
4.根據實施方式2-3之一所述的方法,其中所述翻譯後修飾(ptm)酶是羊毛硫肽合成酶。
5.根據實施方式2-4之一所述的方法,其中所述翻譯後修飾(ptm)酶是lanb型脫水酶、lanc型環化酶和/或雙功能lanm型酶或多功能lankc或lanl型酶。
6.根據實施方式2-5之一所述的方法,其中所述前導序列是來自lana前體肽的前導序列,或者是衍生自lana前體肽的前導序列,或者是帶有來自lana前體肽的共有基序的前導序列。
7.根據實施方式6所述的方法,其中所述前導序列由lanb型脫水酶、lanc型環化酶、雙功能lanm型酶或多功能lankc或lanl型酶識別。
8.根據實施方式2-7之一所述的方法,其中所述前導序列是nisa或proca前導序列或者具有其共有基序。
9.根據前述實施方式之一所述的方法,其中一種或多種分子內鍵通過環化酶或者在弱鹼性條件下形成。
10.根據前述實施方式之一所述的方法,其中脫水殘基是脫氫丙氨酸(dha)或脫氫氨基丁酸(dhb)。
11.根據前述實施方式之一所述的方法,其中分子內鍵是硫醚-或賴丙氨酸-橋。
12.根據前述實施方式之一所述的方法,其中所述外被蛋白是野生型噬菌體外被蛋白。
13.根據實施方式12所述的方法,其中所述噬菌體外被蛋白是或者源自於野生型外被蛋白piii或野生型外被蛋白pviii。
14.根據前述實施方式之一所述的方法,其中所述環肽經由基因融合或者經由通過一個或多個人工引入的半胱氨酸形成的二硫鍵與所述噬菌體顆粒的外被蛋白的c-端連接。
15.一種能夠在噬菌體顆粒的表面上展示環肽的核酸序列,其中核酸序列編碼:
(a)所述噬菌體顆粒的外被蛋白;
(b)由翻譯後修飾(ptm)酶識別的前導序列;和
(c)前體環肽,
其中編碼前體環肽的核酸位於所述噬菌體顆粒的外被蛋白的c-端,
並且其中所述前體環肽能夠通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成分子內鍵。
16.根據實施方式15所述的核酸序列,其從n-端到c-端具有以下排列:
n-(噬菌體外被蛋白)-(由翻譯後修飾(ptm)酶識別的前導序列)-(前體環肽)-c。
17.根據實施方式15-17所述的核酸序列,其中所述前導序列是來自lana前體肽的前導序列,或者是衍生自lana前體肽的前導序列,或者是帶有來自lana前體肽的共有基序的前導序列。
18.根據實施方式15-17之一所述的核酸序列,其中前體環肽包括至少一個或多個絲氨酸或蘇氨酸和一個或多個半胱氨酸或賴氨酸。
19.一種載體,其包括根據實施方式15-18所述的核酸。
20.一種宿主細胞,其包括根據實施方式15-18所述的核酸或根據實施方式19所述的載體。
21.一種在其表面上展示環肽的噬菌體顆粒,其通過根據實施方式1-14之一所述的方法得到。
22.一種在其表面上展示環肽的噬菌體顆粒,其中所述環肽與所述噬菌體顆粒的外被蛋白的c-端連接,並且其中所述多肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。
23.根據實施方式21或22所述的噬菌體顆粒,其還包括載體,該載體包括一個或多個編碼能夠形成所述環肽的前體環肽的核酸序列。
24.根據實施方式23所述的噬菌體顆粒,其中所述載體是實施方式19的載體。
25.一種實施方式21-24中任一個所述的噬菌體顆粒的多樣收集物,其中所述噬菌體顆粒的每一個展示環肽多樣收集物中的環肽,其中所述環肽包括通過將一個或多個脫水殘基與半胱氨酸或賴氨酸偶聯而形成的分子內鍵。
26.一種獲得具有所需特性的環肽的方法,其包括:
(a)提供根據實施方式25所述的噬菌體顆粒的多樣收集物;和
(b)篩選所述多樣收集物和/或從所述多樣收集物選擇,以得到至少一種展示具有所需特性的環肽的噬菌體顆粒。
27.根據實施方式26所述的方法,其中所述的所需特性是與所關注的靶標結合。
工作實施例
實施例1:使用xa因子-切割報導基因檢驗的大腸桿菌(e.coli)中可溶性硫醚-橋接肽的異源性表達和修飾狀態的確認
在以下實施例中,所有分子生物學實驗均根據標準方案進行(ausubel等人,1999)。
載體的構建和模型肽(包括xa因子-切割位點的肽)的可溶性表達:
大腸桿菌中硫醚-橋接羊毛硫抗生素的異源性表達最近已得以說明,其通過前體肽與相關修飾肽一起的共表達實現(shi等人,jamchemsoc.2011mar2;133(8):2338-41)。我們開發出能夠對其在複雜生物樣品中的修飾狀態進行迅速評價並且僅需要微量材料的報導基因肽。這些肽由與人工中心肽融合的前導肽(例如,源自nisa、proca和其他)組成,其包括側面被用於酶促硫醚-橋部分的底物殘基相接的xa因子蛋白酶切割位點和兩個用於檢測的親合力標記(例如his6-和flag-標記)。
將編碼這些肽的dna序列在iptg-誘導型lac-啟動子的控制下克隆到具有氨苄青黴素抗性基因和cole1複製源的大腸桿菌表達質粒中。將產生的質粒與相容性第二質粒(氯黴素抗性,rsf1030複製源)合併,所述相容性第二質粒編碼用於在iptg誘導的pl1aco1-啟動子的控制下進行翻譯後修飾(例如nisb/nisc、procm和其他)的相關酶(羊毛硫肽合成酶)並在相同的大腸桿菌株細胞中共保持。在大腸桿菌株中iptg共誘導時,產生線性前體肽,隨後對其進行酶介導的ptm修飾,其導致底物殘基在側翼與xa因子-切割位點共價連接,並形成穩定的硫醚-橋。
表達之後,將用xa因子蛋白酶處理肽導致識別位點的精氨酸與相鄰殘基之間的肽鍵水解。但是,並沒有得到兩種分別含有切割位點側翼的親合力標記之一的單獨多肽,穩定的硫醚-橋保持了包括兩個親合力標籤的兩種多肽的連接。
因此,在夾心式酶聯免疫吸附測定(elisa)中在xa因子切割之後對兩種親合力標記的共檢測可以因此用於確認模型肽的所需修飾(環化),而對於未修飾的肽(線性),兩個標記中僅有一種可檢測。
對這種xa因子-切割報導基因檢驗在檢測模型肽修飾狀態中的可靠性進行了如下說明(圖1)。含有與nisa-前導肽融合的ompa信號序列、和由位於aswiegrwcn(seq-id.:1)序列(s和c是酶介導的硫醚-橋部分的底物;iegr:xa因子識別基序)側翼的his6-和flag-標記組成的中心肽的模型肽,在存在或不存在nisb和nisc修飾酶共表達的情況下在大腸桿菌mc1061f'中表達。
最為對照修飾,在存在或不存在nisb和nisc的情況下表達其他序列相同的無能力突變肽(s突變成a,或者c突變成a)。
所有菌株均在37℃、220rpm條件下生長,直至早期對數期,通過加入0.25mmiptg誘導肽和酶的表達,培養物生長在22℃下持續過夜。建立起細胞溶解產物,將每個菌株的可溶性蛋白質部分分為四份轉移至抗-hisigg塗覆的384孔板,以經由his6-標記捕獲表達的模型肽。洗滌(4xtbs,lxxa因子反應緩衝液)之後,每個樣品的複製之一用500nmxa因子蛋白酶消化過夜,而另一份不予處理。將板用tbst洗滌之後,加入生物醯化的抗-flagigg,使用鏈黴親和素:sulfo-標記綴合物(使用mesoscalediscoverysectorimager6000測量電化學發光)檢測完整的肽。將不經xa因子處理得到的信號設定成100%(輸入),計算xa因子處理之後相同樣品的信號比(xa-處理之後保留的信號[%])。因此,得到的值反映了xa-切割抗性肽的百分比,其為硫醚-橋形成效率的直接度量。
如圖1所示,在nisb和nisc酶共表達時,得到大約60%切割抗性肽,而在不存在修飾酶的情況下產生的肽幾乎完全切割(左圖)。相反地,含有s至a(中圖)或c至a(右圖)突變的肽變體不是硫醚-橋酶形成的底物,當在nisb和nisc共表達的存在下產生時,完全被xa因子切割。
實施例2:含有nisa-前導的前體肽與piii的c-端融合支持生產細胞中硫醚-橋的酶形成和噬菌體上的展示
在大多數噬菌體展示應用中,所關注的蛋白質或肽與g3p(次要外被蛋白piii)或g8p(主要外被蛋白pviii)的n-端基因融合,分別導致單價和多價展示。piii和pviii的n-端被導向遠離噬菌體顆粒體,據信其支持展示的蛋白質或肽與假定的所關注配體相結合的可及性。
但是,除廣泛應用的所關注的蛋白在n-端上的展示以外,極少有對在piii和pviii對應的c-端上的噬菌體展示的例子(fuh等人,febslett.2000sepl;480(2-3):231-4;held等人,jmolbiol.2004jul9;340(3):587-97)的描述。
對於環狀模型肽在噬菌體上的展示,將含有圖1所示的模型前體肽序列的相同nisa-前導序列克隆到噬菌粒載體中,作為噬菌體g3pn-端和c-端的基因融合物(n-端截短的變體piii-ct),以測試酶法肽修飾和與噬菌體顆粒上展示的相容性。
我們在存在或不存在關聯修飾肽共表達的情況下產生了編碼這些融合物的噬菌體顆粒,並對其進行xa因子-切割報導基因檢驗。
簡言之,在存在或不存在編碼修飾nisb和nisc酶的第二表達質粒的存在下,將用於n-端或c-端模型前體肽(含有nisa-前導序列)與piii的融合物的帶有表達質粒的大腸桿菌mc1061f'細胞分三份生長。將培養物在37℃、220rpm下在24孔板中生長至早期對數期,並在~10個的感染重數下用vcsm13輔助噬菌體感染。通過離心收穫感染細胞之後,通過含有iptg的培養基誘導piii-融合物和修飾酶的表達,使噬菌體產生在22℃下進行16h。通過離心除去生產細胞之後,將含有噬菌體的上清液分四份轉移至塗有抗-m13(santacruz)igg的384孔板中,以經由主要外被蛋白pviii捕獲噬菌體顆粒。
洗滌(4xtbs,lxxa因子反應緩衝液)之後,樣品雙份之一用500nmxa因子蛋白酶消化過夜,而另一份不予處理。
將板用tbst洗滌之後,加入生物素化抗-flagigg(sigma;對於n-端piii-融合物)或生物素化抗-hisigg(r&dsystems;對於c-端piii-融合物),使用鏈黴親和素:sulfo-tag綴合物(電化學發光使用mesoscalediscoverysectorimager6000測量)檢測完整的肽。將不經xa因子處理得到的信號設定成100%(輸入),計算xa因子處理之後相同樣品的信號比(xa-處理之後保留的信號[%])。因此,得到的值反映了xa-切割抗性肽的百分比,其為硫醚-橋形成效率的直接度量。
如圖2a所示,噬菌體產生過程中nisb和nisc酶的共表達導致與piiic-端融合的前體肽的硫醚-橋酶修飾,以及隨後的環肽在噬菌體表面上的展示(左圖)。相反地,相同前體肽與piiin-端的融合物不產生nisb/nisc-介導的修飾,噬菌體上檢測不到環肽(右圖)。這一結果進一步通過使用可溶性表達的麥芽糖-結合蛋白質(mbp)作為前體肽融合載體得到的結果支持,其中c-端的肽融合物被酶修飾(圖2c,左圖),而nisb/nisc不能修飾n-端融合物(圖2c,右圖;樣品製備和檢驗設置如實施例1所述)。在c-端前體肽與piii融合物的上下文中,殘基的突變涉及到硫醚-橋的形成,例如,絲氨酸至丙氨酸(piii-nisa-pep_a/c;圖2b,左圖)或半胱氨酸至丙氨酸(piii-nisa-pep_s/a;圖2b,右圖)消除了環肽的形成和展示,這進一步強調了定製的肽修飾實現的精度。
實施例3:翻譯後修飾肽的c-端piii噬菌體展示是廣泛適用的,並可以轉移至其他酶系統
所述在噬菌體結構蛋白質的c-端上展示翻譯後修飾肽的方法可以很容易地適用於其他酶或半酶修飾系統,如下文概述。在另一實施例中,將模型前體肽與piii的c-端融合(質粒:pl3c_p3.3_mut10-his),其含有prochlorosin3.3(proca3.3)前導肽和核心肽序列gdagioavlaswiegrecnaaagp(seq-id.:6;xa因子切割位點和用於翻譯後修飾的側翼s和c殘基加以下劃線),之後是his6-標記。除對照構建物以外,還將親本核心肽的絲氨酸突變成蘇氨酸(以測試經由脫氫氨基丁酸的甲基羊毛硫氨酸形成),或者將半胱氨酸突變成丙氨酸(以防止硫醚-橋的酶形成)。
將已知修飾若干天然存在的prochlorosin(包括proca3.3)前體的編碼雙功能脫水酶/環化酶的來自原綠球藻mit9313的procm基因由染色體dna擴增,並克隆到大腸桿菌表達質粒中。在存在或不存在procm酶共表達的情況下產生展示模型前體肽的噬菌體,並如上所述進行xa因子-切割報導基因檢驗。當在雙特異性修飾procm酶的存在下產生時,檢測到xa-切割抗性肽融合物具有含有絲氨酸/半胱氨酸-(proca-pep_s/c-piii;圖3,左圖)和蘇氨酸/半胱氨酸-(proca-pep_t/c-piii;圖3,中圖)的核心序列的變體,其分別表明包括羊毛硫氨酸和甲基羊毛硫氨酸的環肽的形成和展示。再次,當參與形成硫醚-橋的半胱氨酸被突變成丙氨酸時,環肽的形成/展示被取消(proca-pep_s/a-piii;圖3,右圖)。
實施例4:環大小靈活的翻譯後修飾的肽的c-端piii噬菌體展示
通過調節酶修飾要求的絲氨酸和蘇氨酸殘基相對於半胱氨酸的間隔,可以在噬菌體上展示環大小合適的c-端piii-肽融合物。在另一實施方式中,建立絲氨酸與半胱氨酸間隔增加的編碼模型前體肽序列,並與piii的c-端融合。在此,將flag-標記和aswiegrecn-(seq-id.:11)、aswaaiegraecn-(seq-id.:12)、aswaaaiegraaaecn-(seq-id.:13)或aswaagaaiegraagaaecn-基序(seq-id.:14;分別能夠實現環大小i,i+7、i,i+10、i,i+13和i,i+17;xa因子切割位點加下劃線)之後以及his6-標記之後的nisa-前導肽與piii的c-端融合。類似地,將ha-標記和aswiegrecn-(seq-id.:11)、aswaaiegraecn-(seq-id.:12)、aswaaaiegraaaecn-(seq-id.:13)或aswaagaaiegraagaaecn-基序(seq-id.:14)之後,以及his6-標記之後的proca3.3-前導肽與piii的c-端融合。分別在存在或不存在關聯修飾nisb/nisc或procm酶的情況下,產生具有nisa-或proca-前導序列的展示模型前體肽的噬菌體,並如上所述進行xa因子-切割報導基因檢驗。使用nisb/nisc(圖5a)和procm(圖5b)酶系統觀察噬菌體上展示的xa因子-切割抗性(硫醚-橋接)肽融合物。如通過切割抗性肽融合物百分比判斷,所測試肽的修飾效率大大獨立於環的大小。
工作實施例說明了與噬菌體piiic端融合的含有前導序列的肽在大腸桿菌中的酶促翻譯後修飾,以及隨後在噬菌體顆粒上的展示。對於與噬菌體主要外被蛋白pviii的c-端前體肽融合物,預期具有類似的結果。
本文所述方法廣泛的應用性很容易是顯而易見的,因為源自於關係甚遠的物種例如乳酸-和藍藻細菌的不同的酶機制/前導肽對均可以有效地用於安裝翻譯後修飾。類似地,該概念可以擴展至其他功能等同的前體肽/酶系統。
總之,經證實,通過脫水酶(例如nisb或雙功能酶例如procm)將絲氨酸和蘇氨酸殘基分別翻譯後脫水成具有高度化學反應性的脫氫丙氨酸和脫氫氨基丁酸,以及通過環化酶(例如nisc或雙功能酶例如procm)形成環肽,與噬菌體展示是相容的。
另外可選地,在弱鹼性條件下,含有脫氫丙氨酸的噬菌體展示的肽可以很容易地與相鄰的半胱氨酸或賴氨酸殘基反應,分別形成硫醚-和賴丙氨酸-橋,以形成許多種在噬菌體顆粒上展示的受限多肽結構。
序列表
莫佛塞斯公司
在噬菌體顆粒上展示環肽的新方法
ms225/pct
ep14199588.6
2014-12-22
14
patentin版本3.5
1
10
prt
人工序列
人工序列
1
alasertrpilegluglyargtrpcysasn
1510
2
424
prt
人工序列
人工序列
2
metlyslysleuleuphealaileproleuvalvalprophetyrser
151015
hisseralagluthrvalglusercysleualalysprohisthrglu
202530
asnserphethrasnvaltrplysaspasplysthrleuaspargtyr
354045
alaasntyrgluglycysleutrpasnalathrglyvalvalvalcys
505560
thrglyaspgluthrglncystyrglythrtrpvalproileglyleu
65707580
alaileprogluasngluglyglyglysergluglyglyglyserglu
859095
glyglyglysergluglyglyglythrlysproproglutyrglyasp
100105110
thrproileproglytyrthrtyrileasnproleuaspglythrtyr
115120125
proproglythrgluglnasnproalaasnproasnproserleuglu
130135140
gluserglnproleuasnthrphemetpheglnasnasnargphearg
145150155160
asnargglnglyalaleuthrvaltyrthrglythrvalthrglngly
165170175
thraspprovallysthrtyrtyrglntyrthrprovalserserlys
180185190
alamettyraspalatyrtrpasnglylyspheargaspcysalaphe
195200205
hisserglypheasngluaspprophevalcysglutyrglnglygln
210215220
serseraspleuproglnproprovalasnalaglyglyglysergly
225230235240
glyglyserglyglyglysergluglyglyglysergluglyglygly
245250255
sergluglyglyglysergluglyglyglyserglyglyglysergly
260265270
serglyasppheasptyrglulysmetalaasnalaasnlysglyala
275280285
metthrgluasnalaaspgluasnalaleuglnseraspalalysgly
290295300
lysleuaspservalalathrasptyrglyalaalaileaspglyphe
305310315320
ileglyaspvalserglyleualaasnglyasnglyalathrglyasp
325330335
phealaglyserasnserglnmetalaglnvalglyaspglyaspasn
340345350
serproleumetasnasnpheargglntyrleuproserleuprogln
355360365
servalglucysargprophevalpheseralaglylysprotyrglu
370375380
pheserileaspcysasplysileasnleupheargglyvalpheala
385390395400
pheleuleutyrvalalathrphemettyrvalpheserthrpheala
405410415
asnileleuargasnlysgluser
420
3
73
prt
人工序列
人工序列
3
metlyslysserleuvalleulysalaservalalavalalathrleu
151015
valprometleuserphealaalagluglyaspaspproalalysala
202530
alapheasnserleuglnalaseralathrglutyrileglytyrala
354045
trpalametvalvalvalilevalglyalathrileglyilelysleu
505560
phelyslysphethrserlysalaser
6570
4
6
prt
人工序列
人工序列
4
leuargasnlysgluser
15
5
5
prt
人工序列
人工序列
5
thrserlysalaser
15
6
24
prt
人工序列
人工序列
6
glyaspalaglyileglnalavalleualasertrpilegluglyarg
151015
glucysasnalaalaalaglypro
20
7
10
prt
人工序列
人工序列
7
leuglugluvalsergluglngluleuasp
1510
8
6
prt
人工序列
人工序列
8
leupheaspleuaspleu
15
9
5
prt
人工序列
人工序列
9
pheasnleuaspval
15
10
8
prt
人工序列
人工序列
10
ileleugluleuglnasnleuasp
15
11
10
prt
人工序列
人工序列
11
alasertrpilegluglyargglucysasn
1510
12
13
prt
人工序列
人工序列
12
alasertrpalaalailegluglyargalaglucysasn
1510
13
16
prt
人工序列
人工序列
13
alasertrpalaalaalailegluglyargalaalaalaglucysasn
151015
14
20
prt
人工序列
人工序列
14
alasertrpalaalaglyalaalailegluglyargalaalaglyala
151015
alaglucysasn
20