棉纖維顏色相關基因及其在調控棉纖維顏色中的應用的製作方法
2023-04-27 22:56:41 2
本發明涉及植物學領域,更具體地涉及棉纖維顏色相關基因及其調控棉纖維顏色中的應用。
背景技術:
棉花是世界上最主要的農作物之一,產量大、生產成本低,使棉製品價格比較低廉。棉纖維能製成多種規格的織物,從輕盈透明的巴裡紗到厚實的帆布和厚平絨,適於製作各類衣服、家具布和工業用布。棉織物堅牢耐磨,能夠洗滌和在高溫下熨燙,棉布由於吸溼和脫溼快速而使穿著舒服,應用廣泛。我國是世界上棉花產量最高的國家之一。
天然彩色棉花(簡稱「彩棉」)是一種棉纖維自身具有棕、綠等不同天然色彩的棉花的統稱。由於其本身帶有天然的顏色,在棉花紡織生產工業上,可以免去漂白、消毒、染色等過程,減輕了紡織過程的汙染,成為真正意義上的綠色產品。天然彩色棉不但可以大大提高紡織工業的經濟效益和社會效益,而且隨著科學技術的迅速發展和人民生活需求、環保意識、健康觀念的不斷提高,彩色棉將成為一個新興產業,為彩色棉育種的深入研究和產業化的實施創造良好的環境。然而,目前彩色棉纖維色素的形成機制還不清楚,通過基因工程手段來調控棉花顏色的研究還有很多不足,本領域迫切需要開發調控彩色棉纖維色素形成的新方法。
技術實現要素:
本發明的目的在於提供一種棉纖維顏色相關基因及其在調控棉纖維顏色中的應用。
在本發明的第一方面,提供了一種棉基因或其編碼蛋白的用途,其特徵在於,所述的棉基因或其編碼蛋白用於選自下組的一種或多種用途:
(a)用於製備促進棉纖維顏色變化的試劑或組合物;
(b)用於調控棉屬植物的棉纖維顏色;
(c)用作鑑別棉纖維顏色的標誌物;
(d)用於製備鑑別棉屬植物的棉纖維顏色的試劑或試劑盒;
(e)用於改良棉屬植物的棉纖維顏色性狀;
其中,所述的棉基因選自下組a:at3gt基因、ghchi基因、或其組合。
在另一優選例中,所述的棉基因還包括選自下組b的基因:ghpal基因、ghc4h基因、ghchs基因、ghdfr基因、ghf3h基因、ghf3′h基因、ghf3'5′h基因、ghans基因、ghccr基因、gh3gt基因、ghhct基因、ghccoaomt基因、或其組合。
在另一優選例中,所述的棉基因包括野生型和突變型。
在另一優選例中,所述的突變型包括突變後編碼蛋白的功能未發生改變的突變形式(即功能與野生型編碼蛋白相同或基本相同)。
在另一優選例中,所述的棉基因包括與野生型基因相比,同源性≥80%(較佳地≥90%,更佳地≥95%)的多核苷酸,只要其編碼的多肽與野生型基因所編碼的蛋白相同或基本相同。
在另一優選例中,所述的棉屬植物為棉花。
在另一優選例中,所述的棉基因具有調控棉纖維顏色的功能。
在另一優選例中,所述的棉基因(即棉纖維顏色相關基因)選自下組:
(i)at3gt基因、ghchi基因、ghpal基因、ghc4h基因、ghchs基因、ghdfr基因、ghf3h基因、ghf3'h基因、ghf3'5′h基因、ghans基因、ghccr基因、gh3gt基因、ghhct基因、ghccoaomt基因、或其組合;
(ii)核苷酸序列與(i)所示的任一基因的同源性≥80%(較佳地≥90%,更佳地≥95%)的多核苷酸;和
(iii)在(i)所示的任一基因的5'端和/或3'端截短或添加1-60個(較佳地1-30,更佳地1-10個)核苷酸的多核苷酸。
在另一優選例中,所述的基因包括dna、cdna、和/或mrna。
在另一優選例中,所述的at3gt基因的cds序列如seqidno.:1所示。
在另一優選例中,所述的at3gt基因的編碼蛋白如seqidno.:2所示。
在另一優選例中,所述的at3gt基因的基因組序列如seqidno.:3所示。
在另一優選例中,所述的ghchi基因包括ghchi-1基因和ghchi-2基因。
在另一優選例中,所述的ghchi-1基因的cds序列如seqidno.:4所示。
在另一優選例中,所述的ghchi-1基因的編碼蛋白如seqidno.:5所示。
在另一優選例中,所述的ghchi-2基因的cds序列如seqidno.:6所示。
在另一優選例中,所述的ghchi-2基因的編碼蛋白如seqidno.:7所示。
在另一優選例中,所述的棉纖維為棉屬(gossypium)植物的種籽纖維。
在另一優選例中,所述的棉纖維顏色包括綠色、棕色、或白色。
在另一優選例中,所述的調控棉纖維顏色包括調控棉纖維顏色為綠色、棕色、和/或白色。
在另一優選例中,所述的調控棉纖維顏色包括調控棉纖維色素的形成。
在本發明的第二方面,提供了一種改良棉屬植物性狀的方法,其特徵在於,包括步驟:
(a)將外源的構建物導入植物細胞,其中所述的構建物含有外源的棉基因序列、促進棉基因表達的外源核苷酸序列、或抑制棉基因表達的外源核苷酸序列,從而獲得導入外源構建物的植物細胞;
(b)將上一步驟獲得的所述導入外源構建物的植物細胞,再生成植株:和
(c)任選地對所述再生的植株進行性狀鑑定,從而獲得性狀改良的植物,
其中,所述植物性狀包括棉纖維的顏色性狀,
並且所述的棉基因選自下組a:at3gt基因、ghchi基因、或其組合。
在另一優選例中,所述的棉基因還包括選自下組b的基因:ghpal基因、ghc4h基因、ghchs基因、ghdfr基因、ghf3h基因、ghf3′h基因、ghf3'5′h基因、ghans基因、ghccr基因、gh3gt基因、ghhct基因、ghccoaomt基因、或其組合。
在另一優選例中,所述的外源核苷酸序列包括幹擾所述棉基因表達的核苷酸序列。
在另一優選例中,所述的外源核苷酸序列包括irna幹擾序列。
在另一優選例中,所述性狀改良包括改良棉纖維的顏色。
在另一優選例中,所述性狀改良包括改良棉纖維顏色為綠色、棕色。
在另一優選例中,所述外源的棉基因序列還包含與orf序列操作性連接的啟動子。
在另一優選例中,所述的啟動子選自下組:組成型啟動子、組織特異性啟動子、誘導型啟動子、或者強啟動子。
在本發明的第三方面,提供了一種促進棉屬植物的棉纖維的綠色性狀的方法,其特徵在於,所述方法包括以下步驟:在所述棉屬植物中,促進at3gt基因的表達或促進at3gt蛋白的活性。
在另一優選例中,所述的棉屬植物包括棕色棉和白色棉。
在另一優選例中,所述的表達為棉纖維組織中的表達。
在另一優選例中,所述的表達為棉纖維組織發育期的表達。
在另一優選例中,所述的棉纖維組織發育期的表達為棉纖維發育第1-30天,較佳地1-20天,更佳地1-15天的表達。
在另一優選例中,所述方法包括給予植物at3gt基因或其編碼的多肽的促進劑。
在另一優選例中,當棉屬植物為棕色棉時,所述方法還包括步驟:抑制ghchi基因的表達。
在另一優選例中,當棉屬植物為白色棉時,所述方法還包括步驟:促進ghchi基因的表達。
在另一優選例中,所述方法包括向植物中導入外源的at3gt基因、或ghchi基因。
在另一優選例中,所述方法包括步驟:
(i)提供一植物或植物細胞;和
(ii)將at3gt基因序列導入所述植物或植物細胞,從而獲得轉基因的植物或植物細胞。
在另一優選例中,所述方法包括步驟:
(a)提供攜帶at3gt基因序列的表達載體的農桿菌;
(b)將植物細胞或組織或器官與步驟(a)中的農桿菌接觸,從而使at3gt的基因序列轉入植物細胞,並且整合到植物細胞的染色體上;
(c)選擇已轉入at3gt基因序列的植物細胞或組織或器官;和
(d)將步驟(c)中的植物細胞或組織或器官再生為植株。
在本發明的第四方面,提供了一種改變棉屬植物的棉纖維顏色變化的方法,所述顏色變化為從棕色變為綠色,其特徵在於,所述方法包括以下步驟:在所述棉屬植物中,抑制ghchi基因的表達或抑制ghchi蛋白的活性。
在另一優選例中,所述方法包括給予棕色棉ghchi基因或其編碼的多肽的拮抗劑。
在另一優選例中,所述的表達為棉纖維組織中的表達。
在另一優選例中,所述的表達為棉纖維組織發育期的表達。
在另一優選例中,所述的棉纖維組織發育期的表達為棉纖維發育第1-30天,較佳地1-20天,更佳地1-15天的表達。
在另一優選例中,所述的拮抗劑包括小分子化合物、irna、sirna、shrna、或其組合。
在本發明的第五方面,提供了一種棉基因或其編碼蛋白的調控劑的用途,其特徵在於,用於調控棉屬植物的棉纖維顏色變化,或用於製備調控棉纖維顏色變化的製劑或組合物。
在另一優選例中,所述的調控劑包括促進劑、抑制劑。
在另一優選例中,所述的組合物包括農用組合物。
在另一優選例中,所述的顏色變化包括從棕色變為綠色、從白色變為綠色、從綠色變為棕色。
在另一優選例中,所述的調控包括促進所述顏色變化或抑制(減緩)所述顏色變化。
在另一優選例中,所述的調控劑包括小分子化合物、或核酸類物質。
在另一優選例中,所述的核酸類物質選自下組:mirna、shrna、sirna、或其組合。
在本發明的第六方面,提供了一種分離的棉纖維顏色相關多肽,其特徵在於,所述的多肽具有調控棉纖維顏色的功能,並且,所述的多肽選自下組:
(i)本發明所述的棉纖維顏色相關多肽;
(ii)將(i)所示的任一多肽經過一個或幾個胺基酸殘基的取代、缺失或添加而形成的,控制棉纖維顏色功能的多肽;
(iii)胺基酸序列與(i)所示的任一多肽的同源性≥95%(較佳地≥98%,更佳地≥99%),具有控制棉纖維顏色功能的多肽。
在另一優選例中,所述的多肽包括:具有seqidno.:2、seqidno.:5、seqidno.:7所示胺基酸序列的多肽。
在本發明的第七方面,提供了一種編碼所述棉纖維顏色相關多肽的基因(即棉纖維顏色相關基因)。
在本發明的第八方面,提供了一種載體,其特徵在於,所述載體含有本發明的第七方面所述的基因。
在本發明的第九方面,提供了一種遺傳工程化的宿主細胞,其特徵在於,所述宿主細胞含有本發明的第八方面所述的載體或基因組中整合有本發明的第七方面所述的基因。
在另一優選例中,所述的宿主細胞選自下組:植物細胞、原核細胞。
在另一優選例中,所述的宿主細胞包括錦葵科植物細胞。
在另一優選例中,所述的宿主細胞包括棉屬植物細胞。
在本發明的第十方面,提供了一種製備本發明的第六方面所述多肽的方法,其特徵在於,所述方法包含:
(a)在適合表達的條件下,培養本發明的第九方面所述的宿主細胞;和
(b)從培養物中分離出本發明的第六方面所述的多肽。
應理解,在本發明範圍內中,本發明的上述各技術特徵和在下文(如實施例)中具體描述的各技術特徵之間都可以互相組合,從而構成新的或優選的技術方案。限於篇幅,在此不再一一累述。
附圖說明
圖1顯示了白色棉、綠色棉和棕色棉的棉纖維實物圖。
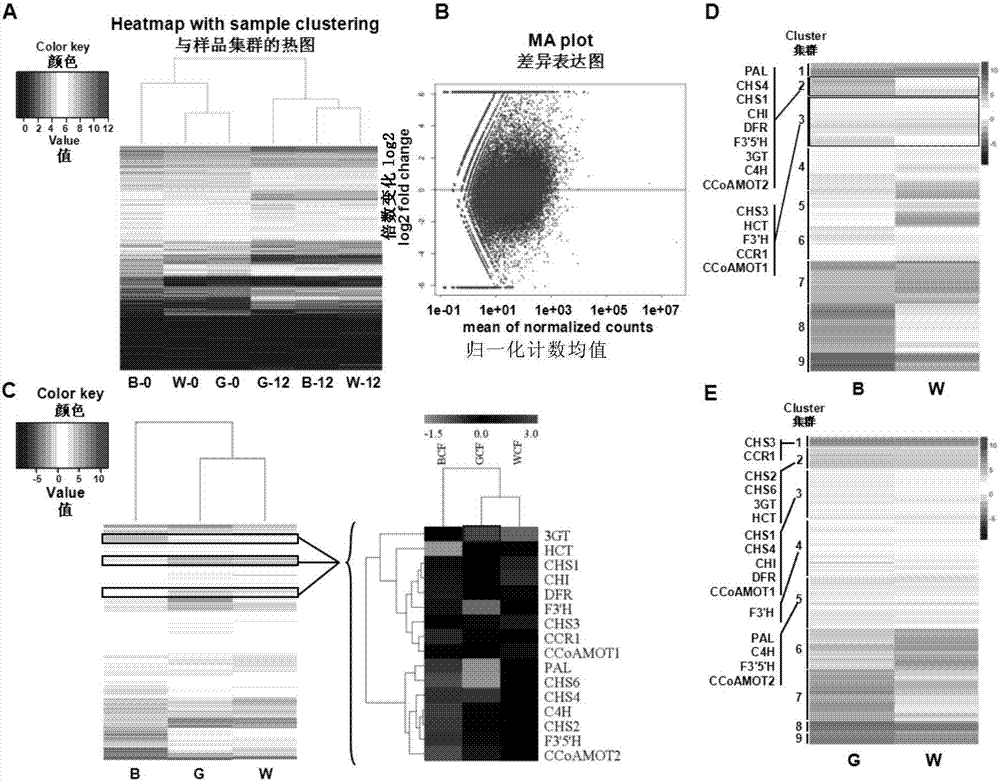
圖2a顯示了白色棉和棕色棉的發育第0天、第12天棉纖維樣品與基因表達的熱圖(b代表棕色纖維brown,w代表白色纖維white,g代表綠色纖維green)。
圖2b顯示了白色棉和棕色棉的發育第0天、第12天棉纖維基因的差異表達分析(紅色點表示差異表達的基因)。
圖2c顯示了白色棉、綠色棉和棕色棉的發育第0天、第12天棉纖維的轉錄譜分析出的苯丙氨酸代謝中的類黃酮生物合成途徑差異表達基因。
圖2d顯示了白色棉和棕色棉在纖維發育12天時的基因差異共表達分析結果,其中顯示了苯丙氨酸途徑中基因集群。
圖2e顯示了白色棉和綠色棉在纖維發育12天時的基因差異共表達分析結果,其中顯示了苯丙氨酸途徑中基因的集群。
圖3a顯示了棕色棉纖維中chi-1和chi-2基因的表達情況。
圖3b顯示了棕色棉纖維中chi-1和chi-2核苷酸與rnai序列比對結果。
圖3c顯示了遺傳轉化棕色棉得到的4個t1代轉化子。
圖3d顯示了1#植株的nptii基因和hpt基因的鑑定結果。
圖3e顯示了1#植株的southernblot鑑定結果。
圖3f顯示了1#植株後代的棉纖維顏色情況。
圖3g顯示了t1代轉化子顏色表現仍為棕色的組1(t2-5#)和表現為白色的組2(t2-24#)和表現為深綠色的(t2-6#)植株的ghchi-1和ghchi-2的表達水平變化。
圖3h顯示了panda-chi-1載體的結構圖。
圖4顯示了1#、2#和3#植株後代的棉纖維顏色情況。
圖4a顯示了2#植株的後代的棉纖維顏色情況。
圖4b顯示了3#植株的後代的棉纖維顏色情況。
圖4c顯示了4#植株的後代的棉纖維顏色情況。
圖5顯示了彩色棉和白色棉纖維質量分析結果。
圖5a顯示了彩色棉(新彩棉1號-7號與白色棉新陸中18號)的纖維長度,強度和馬克隆值的比較分析結果。
圖5b顯示了淺棕色(b3)和深棕色(b4)的纖維質量分析結果。
圖5c顯示了淺綠色(g3)和深綠色(g4)的纖維質量分析結果。
圖5d顯示了淺棕色(b3)和淺綠色(g3)的纖維質量分析結果。
圖5e顯示了深棕色(b4)和深綠色(g4)的纖維質量分析結果。
圖5f顯示了1#轉基因t2代纖維強度的分析結果。
圖5g顯示了1#轉基因t2代纖維馬克隆值的分析結果。
圖5h顯示了1#轉基因t2代纖維長度的分析結果。
圖6a顯示了5#、6#和24#植株的纖維顏色。
圖6b顯示了5#、6#和24#植株的類黃酮代謝途徑的轉錄組分析結果。
圖6c顯示了通過轉擬南芥at3gt基因後,原本顏色為棕色的新彩5棉花纖維,改變為綠色。
圖7a和圖7b顯示了構建的過表達3gt基因的綠色植株的pcr鑑定結果以及2#,3#和4#的轉基因鑑定結果。
圖8顯示了1#轉基因t1代24株的nptii基因轉基因驗證結果和t1代5#,6#和24#的southernblot結果。
具體實施方式
本發明人經過廣泛而深入地研究,首次意外地發現了與棉纖維顏色相關的多種棉基因(包括at3gt基因和ghchi基因等),以及通過促進或抑制所述棉基因的表達來改變棉纖維顏色的方法。以at3gt基因和ghchi基因為例,實驗表明,通過增強at3gt基因的表達,可以調控棉纖維顏色為綠色;通過抑制棕色棉的ghchi基因的表達,可以生成綠色棉後代,在農作物育種中具有應用前景。此外,本發明的所述棉纖維顏色相關基因或蛋白還可用作鑑別棉纖維顏色的標誌物,用於預先或輔助性確定棉屬植物的棉纖維顏色。在此基礎上完成了本發明。
術語
如本文所用,術語「棉基因」、「棉纖維顏色相關基因」、「本發明多核苷酸」、「本發明基因」、「編碼棉纖維顏色相關多肽的基因」可以互換使用,都是指具有調控棉纖維顏色功能的基因。較佳地,本發明基因包括表a所示的基因,尤其是at3gt基因和ghchi基因(包含或不包含其內含子的序列)。
如本文所用,術語「棉纖維顏色相關多肽」、「本發明多肽」、「本發明蛋白」、「棉纖維相關基因編碼的多肽」可以互換使用,都是指來源於棉屬植物的、具有調控棉纖維顏色功能的多肽。較佳地,本發明多肽包括at3gt多肽(seqidno.:2)、ghchi-1多肽(seqidno.:5)、ghchi-2多肽(seqidno.:7),或其衍生的具有相同功能的衍生多肽或活性片段。
如本文所用,「分離的」是指物質從其原始環境中分離出來(如果是天然的物質,原始環境即是天然環境)。如活體細胞內的天然狀態下的多聚核苷酸和多肽是沒有分離純化的,但同樣的多聚核苷酸或多肽如從天然狀態中同存在的其他物質中分開,則為分離純化的。
如本文所用,術語「特異性表達」是指目的基因在植物中特定的時間和/或特定的組織的表達。
如本文所用,「外源的」或「異源的」是指不同來源的兩條或多條核酸或蛋白質序列之間的關係。例如,如果啟動子與目的基因序列的組合通常不是天然存在的,則啟動子對於該目的基因來說是外源的。特定序列對於其所插入的細胞或生物體來說是「外源的」。
棉花
棉花是錦葵科棉屬植物的種子纖維,原產於亞熱帶。植株灌木狀,在熱帶地區栽培可長到6米高,一般為1到2米。花朵乳白色,開花後不久轉成深紅色然後凋謝,留下綠色小型的蒴果,稱為棉鈴。棉鈴內有棉籽,棉籽上的茸毛從棉籽表皮長出,塞滿棉鈴內部。棉鈴成熟時裂開,露出柔軟的棉纖維。常見棉纖維白色至白中帶黃,長約2至4釐米,含纖維素約87~90%。棉花野生種特別是二倍體野生種纖維是棕色或者綠色,在進化和人工馴化過程中出現了白色棉花。
天然彩色棉花(簡稱「彩棉」)是一種棉纖維自身具有棕、綠等不同天然色彩的棉花的統稱。棉纖維為綠色的即為綠色棉,棉纖維為棕色的即為棕色棉。由於彩棉本身帶有天然的顏色,在棉花紡織生產工業上,可以免去漂白、消毒、染色等過程,減輕了紡織過程的汙染,成為真正意義上的綠色產品。
本發明的多核苷酸
在本發明中,相關的棉纖維顏色相關基因包括表a中所示的一種或多種基因。
表a
部分的棉纖維顏色相關基因的序列表信息如下
本發明的多核苷酸可以是dna形式或rna形式。dna形式包括cdna、基因組dna或人工合成的dna。
以at3gt為例,基因組dna可以與seqidno.:3所示的序列相同或者是簡併的變異體。本發明的dna可以是單鏈的或是雙鏈的,dna可以是編碼鏈或非編碼鏈。編碼成熟多肽的編碼區序列可以與seqidno.:1所示的編碼區序列相同或者是簡併的變異體。如本文所用,「簡併的變異體」在本發明中是指編碼具有seqidno.:2的蛋白質,但與seqidno.:1所示的編碼區序列或seqidno.:3所示的基因組序列有差別的核酸序列。對於ghchi-1或其它基因而言,依此類推。
編碼seqidno.:2的成熟多肽的多核苷酸包括:只編碼成熟多肽的編碼序列;成熟多肽的編碼序列和各種附加編碼序列;成熟多肽的編碼序列(和任選的附加編碼序列)以及非編碼序列。
術語「編碼多肽的多核苷酸」可以是包括編碼此多肽的多核苷酸,也可以是還包括附加編碼和/或非編碼序列的多核苷酸。
本發明還涉及上述多核苷酸的變異體,其編碼與本發明有相同的胺基酸序列的多肽或多肽的片段、類似物和衍生物。此多核苷酸的變異體可以是天然發生的等位變異體或非天然發生的變異體。這些核苷酸變異體包括取代變異體、缺失變異體和插入變異體。如本領域所知的,等位變異體是一個多核苷酸的替換形式,它可能是一個或多個核苷酸的取代、缺失或插入,但不會從實質上改變其編碼的多肽的功能。
本發明還涉及與上述的序列雜交且兩個序列之間具有至少50%,較佳地至少70%,更佳地至少80%相同性的多核苷酸。本發明特別涉及在嚴格條件下與本發明所述多核苷酸可雜交的多核苷酸。在本發明中,「嚴格條件」是指:(1)在較低離子強度和較高溫度下的雜交和洗脫,如0.2×ssc,0.1%sds,60℃;或(2)雜交時加有變性劑,如50%(v/v)甲醯胺,0.1%小牛血清/0.1%ficoll,42℃等;或(3)僅在兩條序列之間的相同性至少在90%以上,更好是95%以上時才發生雜交。並且,可雜交的多核苷酸編碼的多肽與seqidno.:2所示的成熟多肽有相同的生物學功能和活性。
本發明還涉及與上述的序列雜交的核酸片段。如本文所用,「核酸片段」的長度至少含15個核苷酸,較好是至少30個核苷酸,更好是至少50個核苷酸,最好是至少100個核苷酸以上。核酸片段可用於核酸的擴增技術(如pcr)以確定和/或分離編碼棉纖維相關多肽的多聚核苷酸。
本發明多肽
本發明的多肽可以是重組多肽、天然多肽、合成多肽,優選重組多肽。本發明的多肽可以是天然純化的產物,或是化學合成的產物,或使用重組技術從原核或真核宿主(例如,細菌、酵母、高等植物、昆蟲和哺乳動物細胞)中產生。根據重組生產方案所用的宿主,本發明的多肽可以是糖基化的,或可以是非糖基化的。本發明的多肽還可包括或不包括起始的甲硫氨酸殘基。
本發明還包括棉纖維顏色相關多肽的片段、衍生物和類似物。如本文所用,術語「片段」、「衍生物」和「類似物」是指基本上保持本發明的天然棉纖維顏色相關多肽相同的生物學功能或活性的多肽。本發明的多肽片段、衍生物或類似物可以是(i)有一個或多個保守或非保守性胺基酸殘基(優選保守性胺基酸殘基)被取代的多肽,而這樣的取代的胺基酸殘基可以是也可以不是由遺傳密碼編碼的,或(ii)在一個或多個胺基酸殘基中具有取代基團的多肽,或(iii)成熟多肽與另一個化合物(比如延長多肽半衰期的化合物,例如聚乙二醇)融合所形成的多肽,或(iv)附加的胺基酸序列融合到此多肽序列而形成的多肽(如前導序列或分泌序列或用來純化此多肽的序列或蛋白原序列,或融合蛋白)。根據本文的教導,這些片段、衍生物和類似物屬於本領域熟練技術人員公知的範圍。
在本發明中,術語「棉纖維顏色相關多肽」指具有棉纖維顏色相關多肽活性的seqidno.:2序列的多肽。該術語還包括具有與棉纖維顏色相關多肽相同功能的、seqidno.:2序列的變異形式。這些變異形式包括(但並不限於):一個或多個(通常為1-50個,較佳地1-30個,更佳地1-20個,最佳地1-10個)胺基酸的缺失、插入和/或取代,以及在c末端和/或n末端添加一個或數個(通常為20個以內,較佳地為10個以內,更佳地為5個以內)胺基酸。例如,在本領域中,用性能相近或相似的胺基酸進行取代時,通常不會改變蛋白質的功能。又比如,在c末端和/或n末端添加一個或數個胺基酸通常也不會改變蛋白質的功能。該術語還包括棉纖維顏色相關多肽的活性片段和活性衍生物。
該多肽的變異形式包括:同源序列、保守性變異體、等位變異體、天然突變體、誘導突變體、在高或低的嚴緊度條件下能與棉纖維顏色相關多肽的dna雜交的dna所編碼的蛋白、以及利用抗棉纖維顏色相關多肽的抗血清獲得的多肽或蛋白。本發明還提供了其他多肽,如包含棉纖維顏色相關多肽或其片段的融合蛋白。除了幾乎全長的多肽外,本發明還包括了棉纖維顏色相關多肽的可溶性片段。通常,該片段具有棉纖維顏色相關多肽序列的至少約10個連續胺基酸,通常至少約30個連續胺基酸,較佳地至少約50個連續胺基酸,更佳地至少約80個連續胺基酸,最佳地至少約100個連續胺基酸。
本發明還提供棉纖維顏色相關多肽或多肽的類似物。這些類似物與天然棉纖維顏色相關多肽的差別可以是胺基酸序列上的差異,也可以是不影響序列的修飾形式上的差異,或者兼而有之。這些多肽包括天然或誘導的遺傳變異體。誘導變異體可以通過各種技術得到,如通過輻射或暴露於誘變劑而產生隨機誘變,還可通過定點誘變法或其他已知分子生物學的技術。類似物還包括具有不同於天然l-胺基酸的殘基(如d-胺基酸)的類似物,以及具有非天然存在的或合成的胺基酸(如β、γ-胺基酸)的類似物。應理解,本發明的多肽並不限於上述例舉的代表性的多肽。
修飾(通常不改變一級結構)形式包括:體內或體外的多肽的化學衍生形式如乙醯化或羧基化。修飾還包括糖基化。修飾形式還包括具有磷酸化胺基酸殘基(如磷酸酪氨酸,磷酸絲氨酸,磷酸蘇氨酸)的序列。還包括被修飾從而提高了其抗蛋白水解性能或優化了溶解性能的多肽。
在本發明中,「棉纖維顏色相關多肽保守性變異多肽」指與seqidno.:2的胺基酸序列相比,有至多10個,較佳地至多8個,更佳地至多5個,最佳地至多3個胺基酸被性質相似或相近的胺基酸所替換而形成多肽。在所述蛋白中,用性能相近或相似的胺基酸進行取代時,通常不會改變蛋白質的功能,在c末端和/或\末端添加一個或數個胺基酸通常也不會改變蛋白質的功能。這些保守性變異多肽最好根據表1進行胺基酸替換而產生。
表1
重組技術和植物改良
本發明棉纖維顏色相關基因的全長序列或其片段通常可以用pcr擴增法、重組法或人工合成的方法獲得。對於pcr擴增法,可根據本發明所公開的有關核苷酸序列,尤其是開放閱讀框序列來設計引物,並用市售的cdna庫或按本領域技術人員已知的常規方法所製備的cdna庫作為模板,擴增而得有關序列。當序列較長時,常常需要進行兩次或多次pcr擴增,然後再將各次擴增出的片段按正確次序拼接在一起。
一旦獲得了有關的序列,就可以用重組法來大批量地獲得有關序列。這通常是將其克隆入載體,再轉入細胞,然後通過常規方法從增殖後的宿主細胞中分離得到有關序列。
此外,還可用人工合成的方法來合成有關序列,尤其是片段長度較短時。通常,通過先合成多個小片段,然後再進行連接可獲得序列很長的片段。
目前,已經可以完全通過化學合成來得到編碼本發明蛋白(或其片段,或其衍生物)的dna序列。然後可將該dna序列引入本領域中已知的各種現有的dna分子(或如載體)和細胞中。此外,還可通過化學合成將突變引入本發明蛋白序列中。
本發明也涉及包含本發明的多核苷酸的載體,以及用本發明的載體或棉纖維顏色相關多肽編碼序列經基因工程產生的宿主細胞,以及經重組技術產生本發明所述多肽的方法。
通過常規的重組dna技術(science,1984;224:1431),可利用本發明的多聚核苷酸序列可用來表達或生產重組的棉纖維顏色相關多肽。一般來說有以下步驟:
(1)用本發明的多核苷酸(或變異體),或用含有該多核苷酸的重組表達載體轉化或轉導合適的宿主細胞;
(2)在合適的培養基中培養的宿主細胞;
(3)從培養基或細胞中分離、純化蛋白質。
本發明的多核苷酸序列可插入到重組表達載體中。術語「重組表達載體」指本領域熟知的細菌質粒、噬菌體、酵母質粒、植物細胞病毒、哺乳動物細胞病毒或其他載體。總之,只要能在宿主體內複製和穩定,任何質粒和載體都可以用。表達載體的一個重要特徵是通常含有複製起點、啟動子、標記基因和翻譯控制元件。
本領域的技術人員熟知的方法能用於構建含本發明多核苷酸和合適的轉錄/翻譯控制信號的表達載體。這些方法包括體外重組dna技術、dna合成技術、體內重組技術等。所述的dna序列可有效連接到表達載體中的適當啟動子上,以指導mrna合成。表達載體還包括翻譯起始用的核糖體結合位點和轉錄終止子。
此外,表達載體優選地包含一個或多個選擇性標記基因,以提供用於選擇轉化的宿主細胞的表型性狀,如真核細胞培養用的二氫葉酸還原酶、新黴素抗性以及綠色螢光蛋白(gfp),或用於大腸桿菌的四環素或氨苄青黴素抗性。
包含上述的適當dna序列以及適當啟動子或者控制序列的載體,可以用於轉化適當的宿主細胞,以使其能夠表達蛋白質。
宿主細胞可以是原核細胞,如細菌細胞;或是低等真核細胞,如酵母細胞;或是高等真核細胞,如植物細胞(如農作物和林業植物的細胞)。代表性例子有:大腸桿菌,鏈黴菌屬、農桿菌;真菌細胞如酵母;植物細胞等。
本發明的多核苷酸在高等真核細胞中表達時,如果在載體中插入增強子序列時將會使轉錄得到增強。增強子是dna的順式作用因子,通常大約有10到300個鹼基對,作用於啟動子以增強基因的轉錄。
本領域一般技術人員都清楚如何選擇適當的載體、啟動子、增強子和宿主細胞。
用重組dna轉化宿主細胞可用本領域技術人員熟知的常規技術進行。當宿主為原核生物如大腸桿菌時,能吸收dna的感受態細胞可在指數生長期後收穫,用cacl2法處理,所用的步驟在本領域眾所周知。另一種方法是使用mgcl2。如果需要,轉化也可用電穿孔的方法進行。當宿主是真核生物,可選用如下的dna轉染方法:磷酸鈣共沉澱法,常規機械方法如顯微注射、電穿孔、脂質體包裝等。
轉化植物也可使用農桿菌轉化或基因槍轉化等方法,例如葉盤法。對於轉化的植物細胞、組織或器官可以用常規方法再生成植株,從而獲得棉纖維顏色變化的植物。
獲得的轉化子可以用常規方法培養,表達本發明的基因所編碼的多肽。根據所用的宿主細胞,培養中所用的培養基可選自各種常規培養基。在適於宿主細胞生長的條件下進行培養。當宿主細胞生長到適當的細胞密度後,用合適的方法(如溫度轉換或化學誘導)誘導選擇的啟動子,將細胞再培養一段時間。
在上面的方法中的重組多肽可在細胞內、或在細胞膜上表達、或分泌到細胞外。如果需要,可利用其物理的、化學的和其它特性通過各種分離方法分離和純化重組的蛋白。這些方法是本領域技術人員所熟知的。這些方法的例子包括但並不限於:常規的復性處理、用蛋白沉澱劑處理(鹽析方法)、離心、滲透破菌、超濾處理、超離心、分子篩層析(凝膠過濾)、吸附層析、離子交換層析、高效液相層析(hplc)和其它各種液相層析技術及這些方法的結合。
重組的棉纖維顏色相關多肽有多方面的用途。例如用於篩選具有調控棉纖維顏色功能的化合物、多肽或其它配體。用表達的重組棉纖維顏色相關多肽篩選多肽庫可用於尋找有價值的能抑制、或促進棉纖維顏色相關多肽功能的多肽分子。
另一方面,本發明還包括對棉纖維顏色相關多肽具有特異性的多克隆抗體和單克隆抗體,尤其是單克隆抗體。本發明不僅包括完整的單克隆或多克隆抗體,而且還包括具有免疫活性的抗體片段、或嵌合抗體。
本發明的抗體可以通過本領域內技術人員已知的各種技術進行製備。例如,純化的棉纖維顏色相關多肽基因產物或者其具有抗原性的片段,可被施用於動物以誘導多克隆抗體的產生。本發明的各類抗體可以利用棉纖維顏色相關基因產物的片段或功能區,通過常規免疫技術獲得。這些片段或功能區可以利用重組方法製備或利用多肽合成儀合成。與棉纖維顏色相關基因產物的未修飾形式結合的抗體可以用原核細胞(例如e.coli)中生產的基因產物來免疫動物而產生;與翻譯後修飾形式結合的抗體(如糖基化或磷酸化的蛋白或多肽),可以用真核細胞(例如酵母或昆蟲細胞)中產生的基因產物來免疫動物而獲得。抗棉纖維顏色相關多肽的抗體可用於檢測樣品中的棉纖維顏色相關多肽。
本發明還涉及定量和定位檢測棉纖維顏色相關多肽水平的測試方法。這些試驗是本領域所熟知的。試驗中所檢測的棉纖維顏色相關多肽水平,可用於解釋棉纖維顏色相關多肽調控棉纖維顏色的功能。
一種檢測樣品中是否存在棉纖維顏色相關多肽的方法是利用棉纖維顏色相關多肽的特異性抗體進行檢測,它包括:將樣品與棉纖維顏色相關多肽特異性抗體接觸;觀察是否形成抗體複合物,形成了抗體複合物就表示樣品中存在棉纖維顏色相關多肽。
本發明的多核苷酸的一部分或全部可作為探針固定在微陣列(microarray)或dna晶片(又稱為「基因晶片」)上,用於分析組織中基因的差異表達分析。用棉纖維顏色相關多肽特異的引物進行rna-聚合酶鏈反應(rt-pcr)體外擴增也可檢測棉纖維顏色相關多肽的轉錄產物。
本發明還提供了一種調控棉屬植物的棉纖維顏色為綠色的方法,包括步驟:促進at3gt基因的表達。在本發明的一個優選例中,所述方法包括步驟:
(a)提供攜帶at3gt基因序列的表達載體的農桿菌;
(b)將植物細胞或組織或器官與步驟(a)中的農桿菌接觸,從而使at3gt的基因序列轉入植物細胞,並且整合到植物細胞的染色體上;
(c)選擇已轉入at3gt基因序列的植物細胞或組織或器官;和
(d)將步驟(c)中的植物細胞或組織或器官再生為植株。
本發明多肽及其編碼序列的應用
本發明提供了棉纖維顏色相關多肽及其編碼序列的用途,用於調控棉纖維的顏色。在本發明中,所述的棉纖維顏色相關多肽及其編碼序列還可用於製備調控棉纖維顏色為綠色的組合物。本發明還涉及棉纖維顏色相關多肽及其編碼序列的促進劑或拮抗劑的用途。
任何可提高at3gt蛋白的活性、提高at3gt蛋白的穩定性、促進at3gt蛋白的表達、延長at3gt蛋白有效作用時間、或促進at3gt的轉錄和翻譯的物質均可用於本發明,作為at3gt的促進劑。所述的促進劑可用於調控棉纖維顏色為綠色。在得知了靶序列後,製備幹擾特定基因表達的幹擾分子的方法是本領域人員熟知的。
本發明的主要優點包括:
(a)提供了棉纖維顏色相關基因及其所編碼的多肽。
(b)提供了一種有針對性地調控棉纖維顏色為綠色的方法。
(c)通過運用類黃酮代謝途徑中的基因改變棉花纖維顏色。
下面結合具體實施例,進一步闡述本發明。應理解,這些實施例僅用於說明本發明而不用於限制本發明的範圍。下列實施例中未註明具體條件的實驗方法,通常按照常規條件,或按照製造廠商所建議的條件。除非另外說明,否則百分比和份數按重量計算。
實施例1
轉錄譜分析彩色棉和白色棉纖維發育過程
分別對白色棉、綠色棉和棕色棉發育第0天、第12天的棉纖維進行轉錄譜分析,結果如圖1所示,從聚類分析上可以看出,0天的樣本聚在一起,12天的樣本聚在一起,並且白色棉和棕色棉的整體基因表達模式是類似的(圖2a),說明實驗是可靠的。
分別對白色棉、綠色棉和棕色棉發育第0天、第12天的棉纖維進行全基因組表達譜分析,結果如圖2b、圖2c所示,在彩色棉,尤其是棕色棉纖維中,苯丙氨酸代謝途徑中的類黃酮途徑眾多基因均表現出明顯的差異。與0天相比,纖維發育12天時,pal,c4h,chs,chi,dfr,f3'h,f3'5'h,3gt等均高表達。
分別對白色棉、綠色棉和棕色棉發育第12天的棉纖維進行基因差異共表達分析,結果如圖2c所示,棕色棉纖維中的類黃酮途徑之中的pal,c4h,chs1,chs2,chs4,chs6,chi都較綠色棉和白色棉纖維中表達水平要高。綠色棉和白色棉表達較為相似,在木質素合成途徑上,尤其是第一步關鍵酶hct的表達水平要比棕色棉纖維要高。並且在棕色棉和白色棉的差異表達基因中,苯丙氨酸代謝途徑中的基因主要被分在了兩個集群中,而綠色棉和白色棉差異的苯丙氨酸代謝途徑中的基因主要被分在了5個集群中,結果如圖2d、圖2e所示。
實施例2
構建幹擾類黃酮代謝途徑關鍵基因chi的載體並遺傳轉化棕色棉
通過實施例1中對棕色棉,綠色棉和白色棉纖維早期發育的表達譜分析表明,棕色棉具有顯著的類黃酮代謝途徑。因此,本實施例通過幹擾pal途徑關鍵基因的表達,研究纖維色素的合成情況。具體地,本實施例選擇chi這個關鍵基因進行幹擾。目前,棉花中有兩個chi基因表達,分別為chi-1和chi-2。chi-1和chi-2均在棕色棉纖維中表達,chi-2的表達水平要高於chi-1(圖3a)。胺基酸比對分析結果表明,chi-1和chi-2有一定的相似度,尤其在酶活性位點上較為保守(圖3b)。本實施例選擇chi-1編碼序列中一段特徵性序列,相應構建幹擾載體的rnai序列(seqidno.:8),其長度為224bp。在cmv35啟動子啟動下,通過1kb的gus作為連接體構成一個反向重複,誘導產生小rna,起到下調基因表達的作用。
通過農桿菌侵染法,遺傳轉化3000朵棉花,t0當代沒有看到纖維顏色發生改變,t1代獲得4個轉化子(圖3c)分別為1#,2#,3#和4#植株。
通過「1#植株」的nptii基因和hpt基因的鑑定,確認1#為轉基因植株(圖3d)。進一步通過southernblot對1#植株進行再鑑定,發現了nptii雜交信號(圖3e),再次確認1#是轉基因植株。1#轉基因植株直接種植的後代共有24株,纖維顏色大致分為3種類型:淺棕色(與受體類似,第1組)、白色(帶有一點點棕色,第2組)和深綠色第3組(見圖3f)。
經鑑定2#,3#和4#植株均是轉基因植株(見圖7)。對1#的t1代植株首先進行了pcr驗證它們都是轉基因(圖8),接著進行ghchi基因的定量檢測,從纖維發育的第0天到第5天,在淺棕色(與受體類似)和深綠色組中,ghchi-1基因的表達水平均比白色(帶有一點點棕色)組中的高,到了第10天,在淺棕色和深綠色組的ghchi-1基因的表達水平均有不同程度降低,在白色組中是略微升高(圖3g)。
對2#,3#和4#植株後代的纖維顏色進行調查,結果如圖4所示,纖維顏色主要分為3種類型,淺棕色、白色和深綠色,與1#轉基因植株後代表現一致。
同時針對部分轉基因樣品進行纖維品質的測定,在陸地棉中,通過比較白色棉,新陸中18號與彩色棉,新彩棉1號-7號的纖維長度,強度和馬克隆值,纖維質量均比彩色棉要好(見圖5a),接著比較了,不同顏色棕色棉(b,代表brown,棕色,b3代表纖維顏色等級,b4顏色比b3深,同理,g,代表green,綠色)(圖5b、圖5c、圖5d、圖5e),發現,顏色越深,纖維長度和強度都變差。同時在轉基因後代中,發現深綠色(第3組)棉纖維在馬克隆值和纖維強度上都表現最差(見圖5f、圖5g、圖5h)。
實施例3
構建過表達3gt基因的載體並遺傳轉化棕色棉
既然類黃酮代謝途徑在棕色棉和白色棉中存在顯著性差異,那麼在轉基因後代中,是否依然存在差異。為研究該問題,通過轉錄組分析比較三種纖維顏色的轉基因後代:5#(b3),6#(g4)和24#(b1)(圖6a),發現類黃酮代謝途徑依舊存在差異(圖6b),不過比野生型的bcf和wcf的差異要小。在這中間,又一次發現gh3gt基因仍然高表達在gcf中,與圖5中的結果保持一致。這就提示3gt可能是在gcf中特異性表達的,調控了綠色的形成。
為研究該問題,在生棕色的(b4)受體「xincaimian5#中過表達擬南芥中的3gt基因(at3gt,genbankaccessionno.at5g17050),意外地發現:通過農桿菌介導的柱頭侵染法,在t0代,有一個棉桃纖維顏色發生了改變,由棕色轉變為綠色(圖6c)。通過pcr鑑定其為轉基因植株(圖7)。說明類黃酮生物合成途徑確實調控了綠色纖維的形成。
在本發明提及的所有文獻都在本申請中引用作為參考,就如同每一篇文獻被單獨引用作為參考那樣。此外應理解,在閱讀了本發明的上述講授內容之後,本領域技術人員可以對本發明作各種改動或修改,這些等價形式同樣落於本申請所附權利要求書所限定的範圍。