一種提高絲狀真菌木質纖維素酶系表達及生物基化學品生產的方法與流程
2023-05-15 04:13:11 2
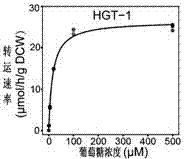
本發明屬於基因工程領域,一種提高真菌降解木質纖維素或是生產纖維素酶、半纖維素酶;提高微生物糖的轉運吸收及其在生物基化學品發酵生產中的應用,涉及2個糖轉運蛋白基因ncu10021(hgt-1)或其同源基因和ncu04963(hgt-2)或其同源基因。
背景技術:
:植物纖維素是自然界中存在最廣泛、含量最豐富的碳水化合物。隨著地球傳統能源的快速消耗、環境汙染和能源危機的日益加劇,纖維素作為潛力巨大、環境友好的可再生能源成為研究的重點。作為地球上含量最豐富的可再生資源,纖維素的利用效率卻遠遠低於社會發展的要求,若能將其高效地轉化為可利用的能源、食料和化學原料等,勢必將會對人類社會的可持續發展起到關鍵作用。要利用這些儲量巨大的生物質資源,需要糖化和發酵兩個基本步驟。其中,如何將生物質高效地水解為單體化合物(葡萄糖、木糖等)已成為其利用過程中的一個迫切需要解決的問題。自然界有一大類真菌能夠自由生長在枯萎或腐爛的植物上,它們是生態圈的分解者。這類真菌大多屬於腐生型的絲狀真菌。絲狀真菌粗糙脈孢菌很早就被發現具有降解利用纖維素的能力,是典型的纖維素降解真菌。在提高生物質降解效率的研究中,傳統誘變技術已經取得了重大進步,提升空間有限。因此,本領域迫切需要開發能夠提高纖維素酶生產從而提供菌株對纖維素利用的技術和方法。絲狀真菌分泌的一系列木質纖維素酶,能將高聚化的纖維素和半纖維素降解為可溶解、可利用的簡單糖類,如葡萄糖、木糖、阿拉伯糖、半乳糖,以及各種寡糖,包括纖維寡糖(二糖、三糖等等)和木寡糖。細胞欲利用這些簡單的糖類,首先需要將這些膜不透過性營養分子轉運進胞內。因此,糖的轉運成為了解析纖維素降解利用的第一步,也是關鍵步驟之一絲狀真菌糖轉運蛋白多屬於膜蛋白家族mfs(majorfacilitatorsuperfamily)。依據轉運的機制,mfs又分為簡單轉運蛋白(uniporter),又稱協助擴散蛋白(facilitateddiffusionprotein),同向轉運蛋白(symporter)以及反向轉運蛋白(antiporter)。uniporter,依靠底物濃度梯度驅動轉運,轉運蛋白主要起著協助運輸的作用。symporter,向同一個方向同時轉運兩種及以上的底物,並以其中一種底物的電化學梯度作為推動力,常見的有sugar/h+,glucose/na+,phosphate/h+,nucleoside/h+以及nitrate/h+等等。antiporter,向反方向協同轉運兩種及以上的底物,驅動力來源和symporter一樣,這一類裡很多都是drag/h+反向轉運蛋白。真菌中的糖轉運蛋白主要是uniporter和symporter/h+類型。由於自然界中脈孢黴能生長在各種乾枯的木質纖維上,可以預見其糖轉運蛋白家族具有很高的功能多樣性。同時,在長期的進化適應過程中,細胞面對不同的生長環境,如酸性-鹼性,碳充足-碳匱乏,會有很好的策略來協同這些轉運蛋白之間的轉運工作,以使得其能高效地吸收外界的營養。通過研究和改造糖轉運蛋白,提高糖的轉運吸收,對改造工程菌(包括酵母,麴黴,木黴)以充分利用培養基中的殘糖發酵,提高糖的轉化利用率具有重要意義。木質纖維素的主要成分是纖維素和半纖維素,其水解產物主要包括葡萄糖、木糖、阿拉伯糖、甘露糖和半乳糖等。實現糖的共利用和高效利用一直是研究熱點。然而,對多數微生物來說,葡萄糖的存在會限制其他糖的利用,稱之為碳代謝物抑制(carboncataboliterepression,ccr)。ccr可以增加微生物對環境的適應和競爭力,所以對微生物自身來說ccr是有利的,而對工業應用來說,則是不利的,優先利用碳源會抑制其他碳源代謝過程所需酶基因的表達,如葡萄糖抑制纖維素酶的表達分泌,從而導致微生物不能利用纖維素。解除ccr的抑制作用,可以實現糖的共利用,快利用,從而提升工業菌株的生產能力。葡萄糖阻遏效應是一個複雜的過程,葡萄糖轉運蛋白是其中重要的參與基因,通過研究調控葡萄糖轉運蛋白,可以調控葡萄糖轉運和下遊阻遏效應,進而促進木糖等其他糖的利用,對實現生物質各類糖的共利用具有重要意義。技術實現要素:本發明的目的是提供一種提高木質纖維素降解及纖維素酶、半纖維素酶表達的方法及其在糖利用中的應用。本發明提供了一種提高絲狀真菌纖維素酶和/或半纖維酶的表達、和/或提高絲狀真菌的纖維素降解活性的方法,包括步驟:抑制絲狀真菌兩個保守的葡萄糖轉運蛋白基因的表達和/或活性,從而提高絲狀真菌纖維素和/或半纖維酶的表達、和/或提高絲狀真菌的纖維素降解活性。在另一優選例中,所述的絲狀真菌包括:脈孢菌(neurospora)、或毀絲黴(myceliophthora)。在另一優選例中,所述的絲狀真菌還包括麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)、或毀絲黴(myceliophthora)。在另一優選例中,所述纖維素降解相關轉運蛋白包括:(a)seqidno.:2、或4所示序列的一種或多種。在另一優選例中,所述的纖維素降解相關轉運蛋白還包括:(b)與seqidno.:2、或2所示序列同源性為50%以上,較佳地為60%以上,更佳地為65%以上的纖維素降解相關轉運蛋白。在另一優選例中,所述抑制包括敲除和/或下調所述的纖維素降解相關轉運蛋白的表達和/或活性。在另一優選例中,所述的纖維素降解相關轉運蛋白至少包括seqidno.:1所示的序列。在另一優選例中,所述的絲狀真菌為脈胞菌,較佳地,為粗糙脈胞菌。本發明還提供了一種纖維素降解相關轉運蛋白或其基因的抑制劑的用途,(a)用於提高絲狀真菌纖維素和/或半纖維酶的表達;和/或(b)用於提高絲狀真菌的纖維素降解活性。在另一優選例中,所述的纖維素降解相關轉運蛋白包括選自(a)seqidno.:2、或4所示序列的一種或多種;或(b)與seqidno.:2、或4所示序列同源性為50%以上,較佳地為60%以上,更佳地為65%以上的纖維素降解相關轉運蛋白。在另一優選例中,所述的纖維素降解相關轉運蛋白的ncbi參考序列號包括:xp_001395794.2、xp_001396930.1、aca66265.1、cac80843.1、caj44796.1、xp_001822181.1、xp_001817192.1、xp_747770.1、xp_746376.1(麴黴);kum58567.1、kum55954.1、xp_002561741.1、eps26451.1(青黴);xp_006964877.1、xp_006966515.1(木黴);eyb25402.1、xp_382817.1(鐮刀黴);xp_003664932.1、xp_003658874.1(毀絲黴)。在另一優選例中,所述的抑制劑包括所述纖維素降解相關轉運蛋白的抗體、抑制性mrna、反義rna、microrna(mirna)、sirna、shrna以及轉運蛋白的活性抑制劑。本發明還提供了一種用於降解纖維素的工程菌,所述工程菌中纖維素降解相關轉運蛋白被敲除或被下調。在另一優選例中,所述的工程菌不表達或基本不表達纖維素降解相關轉運蛋白。在另一優選例中,所述的「基本不表達」指的是與野生型菌株相比,所述工程菌纖維素降解相關轉運蛋白的表達量至少降低了50%,較佳地,至少降低了60%、70%、80%、或90%。在另一優選例中,與野生型菌株相比,所述工程菌纖維素酶和/或半纖維素酶的表達量和/或活性至少提高了20%,較佳地,至少提高了50-80%,更佳地,至少提高了100-500%。在另一優選例中,所述的纖維素降解相關轉運蛋白包括選自seqidno.:2、或4所示序列的一種或多種。在另一優選例中,所述工程菌包括改造自以下野生菌的菌株:脈孢菌(neurospora)、或側孢黴(sporotrichum)、麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)、或毀絲黴(myceliophthora)。本發明還提供了一種生產纖維素酶和/或半纖維素酶的方法,在纖維素原料的存在下,培養所述的工程菌,並從培養物中分離提純所述的纖維素酶和/或半纖維素酶。在另一優選例中,所述的誘導物即纖維素原料包括木質纖維素、結晶纖維素、木聚糖、木質纖維素水解物、木寡糖、纖維寡糖、木糖、或阿拉伯糖。本發明還提供了一種降解纖維素並獲得纖維素降解產物的方法,在木質素原料的存在下,培養所述的工程菌,從而降解木質素並獲得木質素降解產物。本發明還提供了所述工程菌的用途,用於製備纖維素酶和/或半纖維素酶、和/或用於提高纖維素的降解。該方法是利用敲除或突變轉運蛋白基因的微生物菌株降解木質纖維素或是生產纖維素酶、半纖維素酶的方法,所述微生物為脈孢菌(neurospora)、麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)或側孢黴(sporotrichum)。本發明還提供了所述工程菌的用途,用於增強戊糖己糖共利用能力。通過敲除、突變,或降低所述葡萄糖轉運蛋白表達,減低葡萄糖阻遏效應,從而提高混合碳源生長發酵時,戊糖的利用,提高戊糖-己糖共利用能力;本發明還提供了一種增強絲狀真菌葡萄糖轉運吸收和提高生物量生長的方法,通過過表達外源或上調內源所述葡萄糖轉運蛋白基因,可以在葡萄糖或者生物質水解液為碳源,培養工程菌,工程菌葡萄糖轉運吸收能力增強,和/或生物量得到提高;本發明還提供一種增強絲狀真菌生物基化學品發酵能力的方法,其特徵在於,在葡萄糖或者生物質水解液為碳源,培養發明第八方面所述的工程菌,工程菌有機酸或有機醇等生物基化學品發酵能力得到提高;所述轉運蛋白基因包括ncu10021(hgt-1)和ncu04963(hgt-2),敲除其中的任意基因,或突變任意一個轉運蛋白的部分鹼基,或是減弱任意一個轉運蛋白的表達。所述轉運蛋白基因ncu10021的核苷酸序列如序列表中序列1所示,編碼序列表中序列2所示蛋白;所述轉運蛋白基因ncu04963的核苷酸序列如序列表中序列3所示,編碼序列表中序列4所示蛋白。具體的,該方法利用以下任一菌株:以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱轉運蛋白ncu10021基因的菌株;以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱轉運蛋白ncu04963基因的菌株;以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱轉運蛋白ncu10021和ncu04963基因的雙突變體。本發明提供的提高木質纖維素降解和纖維素酶、半纖維素酶表達的方法,是以微生物為出發菌株,敲除其中轉運蛋白基因而得到,或將任意轉運蛋白基因部分鹼基突變而得到,或是減弱任意轉運蛋白基因的表達而得到。所述微生物為脈孢菌(neurospora)、麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)或側孢黴(sporotrichum)。所述敲除或部分鹼基突變或減弱基因表達是針對轉運蛋白基因ncu10021和ncu04963。本發明方法的應用是廣泛的,其為高效表達生產纖維素酶、半纖維素酶或其它蛋白質。其中:生產纖維素酶的方法,是以木質纖維素、結晶纖維素、經過預處理的木質纖維素、纖維寡糖為誘導物,發酵所述的方法得到纖維素酶。生產半纖維素酶的方法,是以木質纖維素、木聚糖、經過預處理的木質纖維素、阿拉伯糖、木寡糖、木糖為誘導物,發酵所述方法得到半纖維素酶。生產其它蛋白的方法,包括將編碼蛋白功能片段的基因導入所述受體菌中形成工程菌,再發酵該工程菌生產蛋白的過程。因此,本發明還涉及這些轉運蛋白基因在構建功能性工程菌中的應用。該應用是將編碼蛋白功能片段的基因導入所述的出發菌株中形成的工程菌。由此形成的高效表達蛋白的工程菌也屬於本發明的技術內容。本發明提供了一種分離的糖轉運蛋白多肽,所述的多肽選自下組:(a)具有seqidno.:4(hgt-2)、或2(hgt-1)中任一條所示胺基酸序列的多肽;(b)將seqidno.:4、或2中任一條所述胺基酸序列的多肽經過一個或幾個胺基酸殘基的取代、缺失或添加而形成的、或是添加信號肽序列後形成的、具有戊糖和/或己糖轉運活性的衍生多肽;(c)序列中含有(a)或(b)中所述多肽序列的衍生多肽;(d)胺基酸序列與seqidno.:4、或2中任意一條所述胺基酸序列的同源性≥85%(較佳地≥90%、更佳地≥95%、98%),並具有戊糖和/或己糖轉運活性的衍生多肽;其中,所述的己糖轉運活性是指將己糖自細胞外轉運至細胞內。在另一優選例中,(d)中所述的衍生多肽來源於以下一種或多種菌株:麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)、毀絲黴(myceliophthora)、球毛殼菌(chaetomiumglobosum)、柄孢黴(podosporaanserina)、稻瘟病菌(magnaportheoryzae)、玉米赤黴(gibberellazeae)。在另一優選例中,所述的細胞為微生物細胞。在另一優選例中,所述的多肽具有將己糖自細胞外轉運至細胞內的活性。在另一優選例中,所述的己糖包括葡萄糖。在另一優選例中,序列如seqidno.:4所示的多肽,具有將葡萄糖自細胞外轉運至細胞內的活性;和/或序列如seqidno.:2所示的多肽,具有將葡萄糖自細胞外轉運至細胞內的活性。本發明提供了一種分離的多核苷酸,所述的多核苷酸為選自下組的序列:(a)編碼本發明第一方面所述多肽的核苷酸序列;(b)編碼seqidno.:4、或2中任一所示多肽的核苷酸序列;(c)如seqidno.:3、或1中任一所示的核苷酸序列;(d)與(a)-(c)任一所述的核苷酸序列互補的核苷酸序列。本發明提供了一種載體,所述的載體含有本發明第七方面所述的多核苷酸。在另一優選例中,所述載體包括表達載體、穿梭載體、整合載體。本發明提供了一種宿主細胞,所述的宿主細胞含有本發明第三方面所述的載體,和/或所述的宿主細胞的染色體整合有外源的本發明第二方面所述的多核苷酸。在另一優選例中,所述的宿主細胞包括真核或原核細胞,優選為真核細胞。在另一優選例中,所述的宿主細胞表達一個或多個本發明第一方面所述的多肽。在另一優選例中,所述的宿主細胞包括酵母(saccharomyces)屬、克魯維酵母菌屬、梭狀芽胞桿菌屬、或絲狀真菌。在另一優選例中,所述的酵母屬包括的釀酒酵母(saccharomycescerevisiae)、摩納酵母(saccharomycesmonacensis)、貝酵母(saccharomycesbayanus)、巴氏酵母(saccharomycespastorianus)、卡氏酵母(saccharomycescarlsbergensis)、裂殖酵母(saccharomycespombe);所述克魯維酵母菌屬(kluyveromyces)包括馬克斯克魯維酵母(kluyveromycesmarxiamus)、乳酸克魯維酵母(kluyveromyceslactis)、脆壁克魯維酵母(kluyveromycesfragilis),樹幹畢赤酵母(pichiastipites)、休哈塔假絲酵母(candidashehatae)、熱帶假絲酵母(candidatropicalis)、運動發酵單孢菌(zymomonasmobilis);所述梭狀芽孢桿菌屬(clostridiumsp.)包括梭熱桿菌(clostridiumthermocellum)、拜氏梭菌(clostridiumbeijerinckii)、丙酮丁醇梭桿菌(clostridiumacetobutylicum)、熱乙酸菌(moorellathermoacetica)、大腸桿菌(escherichiacoli)、產酸克雷伯氏菌(klebsiellaoxytoca)、厭氧桿菌(thermoanaerobacteriumsaccharolyticu)或枯草桿菌(bacillussubtilis);所述絲狀真菌包括嗜熱側孢黴(sporotrichumthermophile)、粗糙脈孢菌(neurosporacrassa)。本發明提供了將所述多肽、多核苷酸、載體或宿主細胞用於將己糖自細胞外轉運至細胞內。本發明還提供了一種促進宿主細胞轉運己糖的方法,包括步驟:在己糖的存在下,培養本發明第九方面所述的宿主細胞。在另一優選例中,當本發明第九方面所述的宿主細胞表達seqidno.:4所示的多肽時,所述的己糖為葡萄糖。在另一優選例中,當本發明第九方面所述的宿主細胞表達seqidno.:2所示的多肽時,所述的己糖為葡萄糖。在另一優選例中,所述的出發菌株包括釀酒酵母,例如釀酒酵母bsw2ap、釀酒酵母eby.vw4000,優選為釀酒酵母bsw2ap菌株本專利的目的在於提供了兩種新的糖轉運蛋白,其具體核苷酸序列及其編碼蛋白的胺基酸殘基序列如下a.葡萄糖轉運蛋白基因hgt-1(ncu10021,來源於粗糙脈胞孢黴,neurosporacrassa)的核苷酸序列如序列表中seqidno.1所示;葡萄糖轉運蛋白基因hgt-1(ncu10021,來源於粗糙脈胞孢黴,neurosporacrassa)編碼蛋白的胺基酸殘基序列如序列表中seqidno.2;b.木糖轉運蛋白基因hgt-2(ncu04963,來源於粗糙脈胞孢黴,neurosporacrassa)的核苷酸序列如序列表中seqidno.3所示;木糖轉運蛋白基因hgt-2(ncu04963,來源於粗糙脈胞孢黴,neurosporacrassa)編碼蛋白的胺基酸殘基序列如序列表中seqidno.4此外,還包含了與以上所述五個轉運蛋白胺基酸殘基序列全長或局部結構域同源度在75%以上的蛋白;該同源度在75%以上的蛋白來自於但不限於以下菌:麴黴(aspergillus)、木黴(trichoderma)、青黴(penicillium)、鐮刀黴(fusarium)、毀絲黴(myceliophthora)、球毛殼菌(chaetomiumglobosum)、柄孢黴(podosporaanserina)、稻瘟病菌(magnaportheoryzae)、玉米赤黴(gibberellazeae)等。所應用的微生物包括但不限於以下菌:酵母屬(saccharomycessp.)的釀酒酵母(saccharomycescerevisiae)、摩納酵母(saccharomycesmonacensis)、貝酵母(saccharomycesbayanus)、巴氏酵母(saccharomycespastorianus)、卡氏酵母(saccharomycescarlsbergensis)、裂殖酵母(saccharomycespombe),克魯維酵母菌屬(kluyveromycessp.)的馬克斯克魯維酵母(kluyveromycesmarxiamus)、乳酸克魯維酵母(kluyveromyceslactis)、脆壁克魯維酵母(kluyveromycesfragilis),樹幹畢赤酵母(pichiastipites)、嗜熱側孢黴(sporotrichumthermophile)、休哈塔假絲酵母(candidashehatae)、熱帶假絲酵母(candidatropicalis)、粗糙脈孢菌(neurosporacrassa)、運動發酵單孢菌(zymomonasmobilis)、梭狀芽孢桿菌(clostridiumsp.)、(clostridiumphytofermentans)、梭熱桿菌(clostridiumthermocellum)、拜氏梭菌(clostridiumbeijerinckii)、丙酮丁醇梭桿菌(clostridiumacetobutylicum)、熱乙酸菌(moorellathermoacetica)、大腸桿菌(escherichiacoli)、產酸克雷伯氏菌(klebsiellaoxytoca)、厭氧桿菌(thermoanaerobacteriumsaccharolyticu)和枯草桿菌(bacillussubtilis)。另一方面,本發明還提供一種使微生物獲得或提高對葡萄糖利用的方法,是將上述轉運蛋白導入微生物細胞(如上述所列)中,所得微生物工程菌株可以將葡萄糖從微生物胞外轉運至胞內,從而提高微生物對葡萄糖的利用效率和相應發酵生產生物基產品的能力。應理解,在本發明範圍內中,本發明的上述各技術特徵和在下文(如實施例)中具體描述的各技術特徵之間都可以互相組合,從而構成新的或優選的技術方案。限於篇幅,在此不再一一累述。附圖說明圖1:wt、ncu10021(△hgt-1)、ncu04963(△hgt-2)突變體在2%微晶纖維素(avicel)上培養7天的產酶動力曲線。圖2:wt、ncu10021與ncu04963雙突變體(△2hgt)在2%微晶纖維素(avicel)上培養7天的產酶動力曲線。圖3:wt、ncu10021與ncu04963雙突變體(△2hgt)在2%微晶纖維素(avicel)上培養5天和7天的纖維素內切酶酶活和木聚糖酶酶活分析。圖4:wt、ncu10021與ncu04963雙突變體(△2hgt)在2%微晶纖維素(avicel)培養基上培養5天和7天後,發酵液上清液中蛋白sds-page分析。圖5:hgt-1轉運蛋白在釀酒酵母中的綠色螢光定位圖。圖6:hgt-1對葡萄糖的同位素轉運的測定。圖7:含有hgt-1的釀酒酵母ehgt-1在葡萄糖平板上的生長。圖8:hgt-2轉運蛋白在釀酒酵母中的綠色螢光定位圖。圖9:hgt-2對葡萄糖的同位素轉運的測定。圖10:含有hgt-2的釀酒酵母ehgt-2在葡萄糖平板上的生長。具體實施方式本發明人經過廣泛而深入的研究,首次意外地發現,絲狀真菌中存在一系列與纖維素降解調控相關的轉運蛋白。實驗證明,當這些轉運蛋白的表達降低或不表達時,絲狀真菌中的纖維素酶或半纖維素酶的表達量會上升,從而能夠提高菌株對木質纖維素的利用,並增強木質纖維素的降解。同時本發明蛋白可以使酵母等微生物以葡萄糖為碳源進行生長。在此基礎上,完成了本發明。葡萄糖轉運蛋白如本文所用,術語「葡萄糖轉運蛋白」、「本發明轉運蛋白」、「本發明轉運蛋白」均指在絲狀真菌中屬於膜蛋白具有調控(半)纖維素酶表達或活性從而與纖維素降解相關的蛋白或其編碼基因。實驗證明,當敲除或下調了本發明轉運蛋白後,能夠有效地提高絲狀真菌中(半)纖維素酶的表達,並進一步提高絲狀真菌對木質纖維素的利用從而加強其降解。優選地,本發明轉運蛋白指的是在脈胞菌中的葡萄糖轉運蛋白,例如,如seqidno.:2、或4所示序列的一種或多種。編碼這些轉運蛋白的核苷酸序列分別如seqidno.:1、或3所示。此外,本發明轉運蛋白還包括來自多種絲狀真菌轉運蛋白。例如,所述的絲狀真菌包括麴黴、木黴、青黴、鐮刀黴或毀絲酶。在這些絲狀真菌的轉運蛋白中,可以找到與本發明seqidno.:2、或4所示序列具有一定同源性的轉運蛋白。本發明人通過篩選發現,在這些絲狀真菌中,具有以下ncbi參考序列號的轉運蛋白與纖維素的降解相關:xp_001395794.2、xp_001396930.1、aca66265.1、cac80843.1、caj44796.1、xp_001822181.1、xp_001817192.1、xp_747770.1、xp_746376.1(麴黴);kum58567.1、kum55954.1、xp_002561741.1、eps26451.1(青黴);xp_006964877.1、xp_006966515.1(木黴);eyb25402.1、xp_382817.1(鐮刀黴);xp_003664932.1、xp_003658874.1(毀絲酶)。優選地,這些轉運蛋白中與本發明seqidno.:2、或4所示序列同源性大於50%,較佳地大於60%,更佳地大於65%的轉運蛋白與纖維素降解有密切的相關性。同時,本領域技術人員可以根據本發明教導在其他絲狀真菌中找到同源性較高的葡萄糖轉運蛋白。此外,本發明人還通過實驗發現,當單個敲除或多個敲除所述的轉運蛋白後,均能夠有效地使(半)纖維素酶活性增加,而不影響菌株的正常生長。纖維素、纖維素酶、半纖維素酶纖維素是自然界中存在最廣泛、最豐富的碳水化合物。多種微生物都具有降解、利用纖維素的能力,其可以分泌多種纖維素水解酶。纖維素酶是一種多酶體系,由多種組份組成。按底物特異性可將纖維素酶分為以下組分:(1)外切β-1,4-葡萄糖酶(exo-beta-1,4-glucanase,ec.3.2.1.91)簡稱為外切酶,又稱為纖維二糖水解酶(cellobiohydrolase)。纖維二糖水解酶可以水解纖維素結晶區,從纖維素鏈的非還原端/還原端開始水解並釋放纖維二糖。(2)內切β-1,4-葡聚糖酶(endo-beta-1,4-glucanase,ec3.2.1.74)簡稱內切酶。纖維素內切酶能夠在纖維素多糖鏈上無定形部分隨機切斷糖苷鍵,產生新的糖鏈末端和長度不一的纖維寡糖。(3)β-1,4-葡萄糖苷酶(beta-1,4-glucosidase,ec3.2.1.21)簡稱纖維二糖酶。纖維二糖水解酶水解可溶性的纖維糊精和纖維二糖生成葡萄糖。(4)其它組分,除了以上三大組分外還有纖維二糖脫氫酶、纖維二糖醌氧化還原酶、磷酸化酶、纖維素酶小體等都會參與纖維素降解過程。半纖維素具有與纖維素一樣的負責結構,其降解需要多種半纖維素酶的共同作用。其中主要有兩種:β-1,4-木聚糖酶和β-木糖苷酶。前者從半纖維素主鏈內部作用於木糖苷鍵,分解成低聚木糖,後者作用於低聚木糖的末端,釋放出單糖。除此之外,降解半纖維素還需要多種側鏈裂解酶的共同作用,主要有乙醯木聚糖酯酶、α-葡萄糖醛酸糖苷酶、α-阿拉伯呋喃糖苷酶等。糖轉運如本文所用,術語「活性多肽」、「本發明多肽及其衍生多肽」、「本發明轉運蛋白」、「己糖轉運蛋白」、「seqidno.:4、或2所示多肽」,均指具有將己糖自細胞外向細胞內轉運活性的hgt-2(seqidno.:4)、或hgt-1(seqidno.:2)所示的多肽及其衍生多肽。如本文所用,術語「己糖」指的是含有6個碳原子的糖類,通常,所述的己糖包括葡萄糖、半乳糖、甘露糖、果糖等。優選地,所述的己糖為葡萄糖等可用於微生物發酵的六碳糖。工程菌本發明「工程菌」、「突變體」可互換使用,均是指將出發菌株中纖維素降解相關轉運蛋白敲除或下調後獲得的具有(增強的)降解纖維素活性的工程菌株。其中,本發明工程菌在改造後,不表達或基本不表達纖維素降解相關轉運蛋白。本領域技術人員應理解,當獲得了轉運蛋白的序列後,可以通過多種常規技術使某出發菌株中的轉運蛋白的表達下調或沉默。通常,可採用同源重組的方法,採用某標記(或抗性)基因置換出發菌株的原始編碼列。當然,還可以採用能夠抑制纖維素降解相關主運蛋白表達和/或活性的其他基因工程手段或物質對該蛋白進行敲除或下調,從而獲得新的轉基因工程菌。這一類物質被稱為「纖維素降解相關轉運蛋白或其基因的抑制劑」或「本發明抑制劑」。例如,所述的抑制劑包括纖維素降解相關轉運蛋白的抗體、抑制性mrna、反義rna、microrna(mirna)、sirna、shrna以及轉運蛋白的活性抑制劑。同樣,在獲得了本發明轉運蛋白的序列或其編碼核苷酸後,本領域技術人員可以根據常規技術手段設計、合成、鑑定並驗證這些抑制劑及其效果。應理解,這些抑制劑可以是完全抑制本發明轉運蛋白的表達和/或活性,也可以是部分抑制,從而獲得(半)纖維素酶活性增強的本發明工程菌。應用通過抑制本發明轉運蛋白,可以人為地提高絲狀真菌中(半)纖維素酶的表達量和/或活性,從而增加菌株對纖維素(尤其是木質纖維素)的利用。例如,當獲得了本發明的工程菌後,可以通過加入纖維素原料的誘導,對所述的工程菌進行培養。此時,所述的菌株會分泌纖維素酶和半纖維素酶,對其進行分離並提純後,就可以獲得較純的纖維素酶和半纖維素酶。由於產生纖維素酶和半纖維素酶的過程中,纖維素原料受其降解,形成降解產物。因此當進一步對培養物進行分離時,就可以獲得纖維素的多種降解產物,例如多種單體化合物:葡萄糖、木糖等。用時利用編碼本發明轉運蛋白的多核苷酸可以製備一種新的重組菌株,所述的菌株可以利用原先具有或不具有己糖轉運能力的出發菌株,通過導入本發明載體,賦予出發菌株新的或更強己糖轉運能力,從而提高碳源的利用率。本發明有益效果本發明通過改造絲狀真菌在纖維素降解的調控因子和糖轉運原件,增強了絲狀真菌對纖維素的利用,提高了生物質降解,增強戊糖利用,提高工程菌種發酵生產纖維素酶和以葡萄糖和生物質及其衍生糖為原料發酵生產各類產品的能力。下面結合具體實施例,進一步闡述本發明。應理解,這些實施例僅用於說明本發明而不用於限制本發明的範圍。下列實施例中未註明具體條件的實驗方法,通常按照常規條件,例如sambrook等人,分子克隆:實驗室手冊(newyork:coldspringharborlaboratorypress,1989)中所述的條件,或按照製造廠商所建議的條件。除非另外說明,否則百分比和份數是重量百分比和重量份數。本發明所採用的原始出發菌株商購於美國真菌菌種保存庫(fgsc),其餘所用材料均為商購。實施例中描述到的各種生物材料的取得途徑僅是提供一種實驗獲取的途徑以達到具體公開的目的,不應成為對本發明生物材料來源的限制。事實上,所用到的生物材料的來源是廣泛的,任何不違反法律和道德倫理能夠獲取的生物材料都可以按照實施例中的提示替換使用。所用引物均由金唯智生物科技有限公司合成。本發明中,纖維素、葡萄糖購自sigma試劑公司;同位素標記的葡萄糖均購自美國radiolabeled化工有限公司。實施例1工程菌的獲得可採用常規技術對出發菌株中的轉運蛋白進行敲除。以粗糙脈胞菌出發菌株(fgsc2489)(購自fgsc)為例,根據同源重組的原理,利用hph基因(潮黴素b抗性基因)置換ncu10021的整個開放閱讀框(openreadingframe,orf)。1)設計同源臂特異性引物上遊同源臂特異性引物:ncu10021-5f:gtaacgccagggttttcccagtcacgacgcccaccccagacccagcact(seqidno.:5);ncu10021-5r:atccacttaacgttactgaaatctccaacacatggtgtttgttgtgactcttgtgt(seqidno.:6)下遊同源臂特異性引物:ncu10021-3f:ctccttcaatatcatcttctgtctccgacagctgcatattttgagctcctcgtg(seqidno.:7);ncu10021-3r:gcggataacaatttcacacaggaaacagccgcacacgtccgcacttcct(seqidno.:8)擴增目標基因上下遊同源臂;然後利用引物從質粒pcsn44中擴增hph抗性元件。將上述擴增所得3個片段利用酵母組裝的方法連接至prs426中。引物如下所示hph-f:gtcggagacagaagatgatattgaaggagc(seqidno.:9)hph-r:gttggagatttcagtaacgttaagtggat(seqidno.:10)2)將上述所獲得質粒作為模板,用seqidno.:5、8所示的引物擴增ncu10021敲除元件,凝膠純化pcr產物。3)將上述pcr產物10µg電擊轉化至野生型菌株(fgsc2489,a),通過含有潮黴素的平板篩選陽性轉化子。4)提取轉化子基因組dna,以其為模板,利用引物擴增,其中hphdc是驗證基因敲除的通用引物,如果可以擴增得到目的片段,則說明這些轉化子發生了預期的同源重組。引物如下所示:ncu10021dc:cgcacacgtccgcacttcct(seqidno.:11)hphdc:tgcaataggtcaggctct(seqidno.:12)5)將所獲得的轉化子與野生型菌株雜交(fgsc4200,a),將雜交得到的子囊孢子熱激(60℃,45min)後,稀釋塗布在平板上,萌發後挑至斜面上,生長7天後,提取基因組,利用pcr進行鑑定。其餘工程菌株均根據類似方法獲得,所需的引物如表1所示:表1引物seqidno.:序列ncu04963-5f:13gtaacgccagggttttcccagtcacgacgtagctacctacctcgagacgncu04963-5r:14atccacttaacgttactgaaatctccaactgtagtacaggaggctttgcncu04963-3f:15ctccttcaatatcatcttctgtctccgacagctgcccatctaaaggaacagtcgncu04963-3r:16gcggataacaatttcacacaggaaacagcgacacagttcgaatagcagcncu04963dc:17gacacagttcgaatagcagc實施例2工程菌的驗證提基因組dna:採用酚氯仿法從孢子中提取提基因組dna1)rcr採用25μl體系,包括12.5μldreamtaqpcrmastermix12.5μl,9.5μl水(nuclease-free),上下遊引物各1μl,基因組dna1μl。2)按如下條件進行pcr擴增95度預變性5分鐘,94度變性30秒,52度退火45秒,72度延伸2分鐘,按同樣條件進行34個循環,最後72度退火10分鐘。3)110v電壓,1%瓊脂糖凝膠電泳30分鐘,凝膠成像系統下看基因擴增條帶。4)保菌:30%甘油保菌後放於-80℃冰箱保種。實施例3單基因敲除的突變體生長纖維素、半纖維素產量測定利用實施例1的方法、實施例2的鑑定,構建了如下菌株並對其進行產酶動力學測定:以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱轉運蛋白基因ncu10021(hgt-1);以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱轉運蛋白基因ncu04963(hgt-2)。轉運蛋白工程菌和野生型粗糙脈孢菌分別在2%結晶纖維素培養基上培養7天,每天取培養基上清,離心後進行一系列驗證試驗。方法:總的產酶測定取20μl上清液,加入1mlbradford蛋白顯色溶液,室溫下反應5min,在595nm處測定吸光值。標準曲線用小牛血清蛋白作為標準蛋白樣繪製。結果:在2%結晶纖維素條件下培養5天和7天,突變株ncu10021培養基上清液中的產酶水平分別提高了40%和53%(圖1)。在2%結晶纖維素條件下培養5天和7天,突變株ncu4963培養基上清液中的產酶水平分別提高了48%和35%(圖1)。實施例4雙基因敲除的突變體生長活性研究以及酶活性測定轉運蛋白工程菌和野生型粗糙脈孢菌分別在2%結晶纖維素培養基上培養7天,每天取培養基上清,離心後進行一系列驗證試驗。利用實施例1的方法、實施例2的鑑定、實施例3的產酶測定,構建了ncu10021和ncu4963雙缺菌株(△2hgt)並進行了產酶測試。方法:4.1.1酶活測定(1)內切-1,4-β-葡聚糖酶活力的測定方法:將粗酶液用0.1m醋酸鈉緩衝液稀釋適宜的倍數,終體積為0.5ml,放入40℃水浴鍋內預熱,取出,加入0.5mlmegazymeazo-cm-cellulose底物溶液,混勻,40℃溫育10min,用2.5ml沉澱溶液終止反應,室溫靜置10min,混勻,1000g離心10min,590nm波長下測od。空白組使用滅活的酶液作對照。(2)內切-1,4-β-木聚糖酶活力的測定方法:將粗酶液用0.1m醋酸鈉緩衝液稀釋適宜的倍數,終體積為0.5ml,放入40℃水浴鍋內預熱,取出,加入0.5mlmegazymeazo-xylan底物溶液,混勻,40℃溫育10min,用2.5ml沉澱溶液終止反應,室溫靜置10min,混勻,1000g離心10min,590nm波長下測od。空白組使用滅活的酶液作對照。(4)活性定義:1ml酶液於40℃下,每min水解底物產生1μmol產物的酶量定義為1個酶活力單位。蛋白質sds-page實驗在2%結晶纖維素avicel培養7天後,離心得到培養基上清液,上樣,做蛋白質電泳。結果:轉運蛋白對纖維素酶、半纖維素酶表達水平的影響在2%結晶纖維素條件下培養5天,雙缺突變株△2hgt培養基上清液中的酶產量比野生型提高了180%(圖2)、纖維素內切酶活力提高了260%(圖3)、木聚糖酶活提高了190%(圖3)。蛋白膠圖顯示與以上數據吻合(圖4)。在2%結晶纖維素條件下培養5天,雙缺突變株△2hgt培養基上清液中的酶產量比野生型提高了180%(圖2)、纖維素內切酶活力提高了230%(圖3)、木聚糖酶活提高了180%(圖3)。蛋白膠圖顯示與以上數據吻合(圖4)。以上結果顯示,以脈孢菌(neurospora)為出發菌株構建的部分鹼基突變或敲除或減弱表達轉運蛋白基因ncu10021和ncu04963,以及兩者同時被部分鹼基突變或敲除或減弱表達,能顯著提升細胞降解纖維素的能力和產纖維素、半纖維素酶的水平。實施例5hgt-1是葡萄糖轉運蛋白,使微生物獲得轉運和利用葡萄糖能力一、hgt-1基因表達載體的構建利用引物hgt-1-f(序列:5』-cggactagtatgtcgtccgcccacgag-3』)(seqidno.:18)和hgt-1-r(序列:5』-ccggaattcaacctgcttctcagcag-3』)(seqidno.:19)從粗糙脈孢菌的cdna中pcr擴增hgt-1基因的編碼閱讀框,pcr反應體系為:5×phusionhfbuffer10µl,10mmdntps1µl,hgt-1-f2.5µl,hgt-1-r2.5µl,cdna1µl,phusiondnapolymerase0.5µl,水32.5µl。pcr反應條件為:先98℃30s;然後98℃10s,65℃30s,72℃1.5min,35個循環;最後72℃10min,4℃10min。pcr反應結束後,使用限制性內切酶spei和ecori將pcr產物和質粒prs426-pgk1[該質粒構建根據參考文獻(galazka,j.m.,etal.,2010.cellodextrintransportinyeastforimprovedbiofuelproduction.science.330,84-86.)],進行雙酶切,並將兩種雙酶切產物進行連接,將連接產物用限制性內切酶進行酶切鑑定,再進行測序,測序結果表明hgt-1基因的核苷酸序列如序列表中序列1所示,表明獲得了序列及插入位置正確的攜帶hgt-1基因的重組表達質粒,命名為prs426-hgt-1。將質粒prs426-hgt-1和prs426-pgk1轉入釀酒酵母eby.vw4000(wieczorke,r.,etal.,1999.concurrentknock-outofatleast20transportergenesisrequiredtoblockuptakeofhexosesinsaccharomycescerevisiae.febslett.464,123-128.)中,分別命名為ehgt-1和eprs426,如圖5所示。二、hgt-1對葡萄糖的轉運的測定1、分別挑取ehgt-1和對照菌株e426單克隆,將其接種在10ml以2%麥芽糖為碳源的sc-ura培養基(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l)中30℃培養過夜(10-12小時)至菌體濃度為1.5-2.0(od600);2、離心收集菌體(4000rpm,5min)後,用冰預冷assaybuffer(100mmtris-citratebufferph5.0)洗三次,重懸至od600為20;3、將菌體分裝至1.5ml離心管(100ul/管)中,其中三個用於烘乾稱其乾重,其餘置於冰上,以備使用。4、反應前將菌體於30攝氏度,放置5min。5、100ul細胞懸液加入50ul不同濃度同位素標記的葡萄糖溶液(糖濃度:500μm,100μm,20μm,5μm,1μm),反應120s後加入1ml冰水終止反應,6、10000rpm離心1min來收集細胞,冰水洗滌兩次,離心,去上清。7、用500ml,0.1mmnaoh重選菌體,轉移至裝有3mlultimagoldscintillatioinfluid小瓶中,測定其放射量,並計算單位細胞乾重單位時間的轉運量。最終結果圖6所示。釀酒酵母eby.vw4000缺失17個己糖轉運蛋白和3個麥芽糖/葡萄糖轉運蛋白,而失去了轉運葡萄糖的能力。由於hgt-1的導入,重組酵母ehgt-1恢復了對葡萄糖的轉運能力,其中如圖6,hgt-1對葡萄糖的親和力km為16.13±0.95μm,最大轉運速率為26.42±0.38μmol/h/gramdcw。三、hgt-1使微生物具有利用葡萄糖的能力1、分別挑取ehgt-1和對照菌株eprs426單克隆,將其接種在10ml以2%麥芽糖為碳源的sc-ura培養基(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l)中30℃培養過夜(10-12小時)至菌體濃度為1.5-2.0;2、離心收集菌體(4000rpm,5min)後,用冰預冷雙蒸水洗三次,重懸至od600為0.3;3、等梯度稀釋菌夜(100,10-1,10-2);4、將等梯度稀釋的酵母點在以2%麥芽糖為碳源的sc-ura平板中(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l,2%瓊脂),置於30℃培養箱中培養。結果如圖7所示,相比對照菌株(eprs426),重組酵母(ehgt-1)快速利用葡萄糖的能力。說明hgt-1的導入,釀酒酵母恢復了以葡萄糖為碳源進行生長的能力。綜合所述,hgt-1對葡萄糖具有良好的轉運能力(26.42±0.38μmol/h/gramdcw)和極高的親和性(km為16.13±0.95μm),並且hgt-1導入釀酒酵母后,可使其恢復利用葡萄糖為碳源的能力。實施例6hgt-2是葡萄糖轉運蛋白,使微生物獲得轉運和利用葡萄糖生長及乙醇發酵能力一、hgt-2基因表達載體的構建利用引物hgt-2-f(序列:5』-ggactagtatgggcttgtcactcaagaag-3』)(seqidno.:20)和hgt-2-r(序列:5』-ccggaattcagctgcctcttcagtgatatc-3』)(seqidno.:21)從粗糙脈孢菌的cdna中pcr擴增hgt-2基因的編碼閱讀框,pcr反應體系為:5×phusionhfbuffer10µl,10mmdntps1µl,hgt-2-f2.5µl,hgt-2-r2.5µl,cdna1µl,phusiondnapolymerase0.5µl,水32.5µl。pcr反應條件為:先98℃30s;然後98℃10s,65℃30s,72℃1.5min,35個循環;最後72℃10min,4℃10min。pcr反應結束後,使用限制性內切酶spei和ecori將pcr產物和質粒prs426-pgk1[該質粒構建根據參考文獻(galazka,j.m.,etal.,2010.cellodextrintransportinyeastforimprovedbiofuelproduction.science.330,84-86.)],進行雙酶切,並將兩種雙酶切產物進行連接,將連接產物用限制性內切酶進行酶切鑑定,再進行測序,測序結果表明hgt-2基因的核苷酸序列如序列表中序列1所示,表明獲得了序列及插入位置正確的攜帶hgt-2基因的重組表達質粒,命名為prs426-hgt-2。將質粒prs426-hgt-2和prs426-pgk1轉入釀酒酵母eby.vw4000(wieczorke,r.,etal.,1999.concurrentknock-outofatleast20transportergenesisrequiredtoblockuptakeofhexosesinsaccharomycescerevisiae.febslett.464,123-128.)中,分別命名為ehgt-2和eprs426,如圖8所示。二、hgt-2對葡萄糖的轉運的測定1、分別挑取ehgt-2和對照菌株eprs426單克隆,將其接種在10ml以2%麥芽糖為碳源的sc-ura培養基(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l)中30℃培養過夜(10-12小時)至菌體濃度為1.5-2.0(od600);2、離心收集菌體(4000rpm,5min)後,用冰預冷assaybuffer(100mmtris-citratebufferph5.0)洗三次,重懸至od600為20;3、將菌體分裝至1.5ml離心管(100ul/管)中,其中三個用於烘乾稱其乾重,其餘置於冰上,以備使用。4、反應前將菌體於30攝氏度,放置5min。5、100ul細胞懸液加入50ul不同濃度同位素標記的葡萄糖溶液(糖濃度:2000μm,500μm,100μm,20μm,5μm,1μm),反應120s後加入1ml冰水終止反應,6、10000rpm離心1min來收集細胞,冰水洗滌兩次,離心,去上清。7、用500ml,0.1mmnaoh重選菌體,轉移至裝有3mlultimagoldscintillatioinfluid小瓶中,測定其放射量,並計算單位細胞乾重單位時間的轉運量。最終結果如圖9所示。釀酒酵母eby.vw4000缺失17個己糖轉運蛋白和3個麥芽糖/葡萄糖轉運蛋白,而失去了轉運葡萄糖的能力。由於hgt-2的導入,重組酵母ehgt-2恢復了對葡萄糖的轉運能力,其中如圖9,hgt-2對葡萄糖的親和力km為98.97±22.02μm,最大轉運速率為78.11±4.46μmol/h/gramdcw。三、hgt-2使微生物具有利用葡萄糖的能力1、分別挑取ehgt-2和對照菌株eprs426單克隆,將其接種在10ml以2%麥芽糖為碳源的sc-ura培養基(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l)中30℃培養過夜(10-12小時)至菌體濃度為1.5-2.0;2、離心收集菌體(4000rpm,5min)後,用冰預冷雙蒸水洗三次,重懸至od600為0.3;3、等梯度稀釋菌夜(100,10-1,10-2);4、將等梯度稀釋的酵母點在以2%麥芽糖為碳源的sc-ura平板中(配方:無氨基酵母氮源6.7g/l,酵母合成缺失培養基補充物1.4g/l,麥芽糖20g/l,亮氨酸,組氨酸和色氨酸各20mg/l,2%瓊脂),置於30℃培養箱中培養。結果如圖10所示,相比對照菌株(eprs426),重組酵母(ehgt-2)快速利用葡萄糖的能力。說明hgt-2的導入,釀酒酵母恢復了以葡萄糖為碳源進行生長的能力。對該工程酵母進行乙醇發酵試驗,對照菌不能利用乙醇發酵,發酵4天後,工程菌乙醇發酵濃度達到5克/升,顯著提高了工程菌乙醇發酵能力;綜合所述,hgt-2對葡萄糖具有良好的轉運能力(78.11±4.46μmol/h/gramdcw)和較高的親和性(km為98.97±22.02μm),並且hgt-2導入釀酒酵母后,可使其恢復利用葡萄糖為碳源的能力。實施例7通過協同表達hgt-2,可以提高嗜熱毀絲黴蘋果酸發酵能力1、hgt-2過表達載體(pan52-hgt)的構建以粗糙脈孢菌fgsc#2489(購自真菌遺傳保藏中心,fungalgeneticsstockcenter)葡萄糖條件下的cdna為模板,利用pcr擴增出葡萄糖轉運蛋白編碼基因hgt-2(ncu04963,序列seqidno.:3)。引物如下:hgt-f:(seqidno.:22)cggactagtatgggcttgtcactcaagaaghgt-r:(seqidno.:23)cgcggatcctcaagctgcctcttcagtg用spei和ecori酶切後,連入用同樣酶雙酶切的線性化質粒pan52-tb-mtpgpda中,從而得到hgt-2表達載體,命名為:pan52-hgt。2、將表達載體(pan52-hgt)導入嗜熱毀絲黴2.1將嗜熱毀絲黴jg207菌株在mm斜面培養基[50×vogel’s鹽20ml,蔗糖20g,瓊脂15g,組氨酸(50mg/ml)20ml,定容體積到1l,高壓滅菌。50×vogel’s鹽(1l):檸檬酸三鈉(1/2h2o)150g,無水kh2po4250g,無水nh4no3100g,mgso47h2o10g,cacl22h2o5g,微量元素鹽溶液5ml,生物素(0.1mg/ml)2.5ml,定容體積到1l]上45℃培養10天後待用。2.2嗜熱毀絲黴原生質體轉化1)菌絲體準備將成熟的嗜熱毀絲黴孢子,用0.05%吐溫-80滅菌水收集,經擦鏡紙過濾除去菌絲後,塗布於鋪有玻璃紙的mm平板,45℃培養14h。2)原生質體製備將帶有菌絲的玻璃紙放置於30ml裂解液(配方:0.15g裂解酶,無菌操作加入30ml溶液a,過濾除菌;溶液a:1.0361g磷酸二氫鉀,21.864g山梨醇,溶於90ml去離子水,氫氧化鉀調ph到5.6,定量至100ml,高溫滅菌)中,28℃裂解2h,每隔20min輕輕搖動。而後經過擦鏡紙過濾後,2000rpm4℃離心10min,棄上清,加入4ml溶液b(0.735g氯化鈣,18.22g山梨醇,1mltris-hcl1mph7.5,溶於90ml去離子水,鹽酸調ph到7.6,定量至100ml,高溫滅菌),2000rpm4℃離心10min;棄上清,按200μl/質粒加入一定體積溶液b。3)原生質體轉化預冷的15ml離心管中,依次加入50μl預冷peg(12.5gpeg6000,0.368g氯化鈣,500μltrishcl1mph7.5。),10μl用hindiii線性化的質粒pan52-hgt,200μl原生質體。放置冰上20min後加入2ml預冷peg溶液,室溫5min,加入4ml溶液b,輕輕混勻。取3ml上述溶液加入12ml融化的含相應抗生素mm培養基中,置於平板中,45℃培養,2d-4d後於體式顯微鏡下挑取單個菌絲體於相應抗性平板生長2.3嗜熱毀絲黴轉化子驗證1)基因組提取採用酚氯仿法從上述轉化過程中挑選的轉化子提取基因組dna,具體包括以下操作:1)2.0ml的無菌的dna提取管中加入200mg的鋯珠及1ml的裂解液(lysisbuffer,配方:0.2mtrishcl(ph7.5),0.5mnacl,10mmedta,1%sds(w/v)),挑取平板中生長的嗜熱毀絲黴菌絲於dna提取管中。2)將所有dna提取管置於助磨器上,最大轉速振蕩30s,重複兩次。3)65℃水浴30min,在水浴過程中每個幾分鐘取出漩渦振蕩。4)水浴結束後取出,每管加入80μlph7.5的1m的trishcl中和。5)加入400μl的酚:氯仿(1:1),13000rpm離心5分鐘。6)取300μl上清液於新的1.5mlep管中,加入600μl95%的乙醇(dna級)。7)冰上孵育一小時,隨後4℃,13000rpm離心,可看到白色的dna沉澱到ep管底部。8)用75%的酒精(dna級)400μl清洗,4度13000rpm離心,輕輕取出上清液。9)將ep管置於真空濃縮儀中,真空乾燥酒精。10)加入50μlddh2o溶解dna,用nanodrop測dna濃度,測完濃度後將提取的dna置於-20℃冰箱保存,以備下一步進行pcr驗證。2)pcr驗證嗜熱毀絲黴轉化子以提取的轉化子基因組dna為模版,用引物hgt-f及hgt-r對轉化子進行基因pcr驗證。pcr反應體系為:5×phusiongcbuffer4μl,10mmdntps0.2μl,引物各1μl,基因組1μl,dmso0.6μl,phusiondnapolymerase0.1μl,去離子水12.1μl。pcr反應條件為:先98℃30s;然後98℃10s,62℃30s,72℃1.5min,30個循環;最後72℃10min,4℃10min。3)對pcr擴增產物進行1%瓊脂糖凝膠電泳(110v電壓,30分鐘),在凝膠成像系統下觀察基因擴增條帶,結果表明經hindiii線性化的pan52-hgt已整合到了嗜熱毀絲黴基因組中。3、嗜熱毀絲黴轉化子生產蘋果酸能力測定將上述驗證的轉化子全部接種至250ml三角瓶中的50ml以葡萄糖為碳源培養基中(配方:碳源75g/l,蛋白腖6.0g/l,0.15g/lkh2po4,0.15g/lk2hpo4,0.10g/lcacl22h2o,0.10g/lmgso47h2o,碳酸鈣80.0g/l,1ml/l0.5g/l生物素,1ml/l微量元素液;微量元素配方(100ml):5gc6h8o7h2o,5gznso47h2o,1gfe(nh4)2(so4)6h2o,0.25gcuso45h2o,0.05gmnso4h2o,0.05gh3bo3,0.05gnamoo42h2o,溶於水中,定容至100ml),接種量為2.5*105個/ml,45℃條件下150rpm培養,第六天取樣測定蘋果酸含量。1)樣品處理:取1ml發酵液於15ml離心管中,並添加1ml1mh2so4,而後80℃下放置30min,每個隔10min進行充分震蕩。之後將2ml雙蒸水添加至離心管中,充分震蕩後,取1ml液體於1.5ml離心管中,12000rpm離心10min,取上清液測定c4-二羧酸含量。2)蘋果酸含量測定處理後的樣品用高效液相色譜測定蘋果酸含量,其中檢測器為紫外檢測器,流動相為5mmh2so4,流速為0.5ml/min。結果顯示第六天時轉化子的蘋果酸產量與對照菌株jg207相比提高20%。實驗說明hgt-2在嗜熱毀絲黴中過表達時,能顯著促進嗜熱毀絲黴利用葡萄糖進行蘋果酸發酵。另外,降低嗜熱毀絲黴內源hgt1/2同源基因表達,也有利於蘋果酸合成,發酵水平可提高10%。實施例8通過協同表達hgt1/hgt-2是葡萄糖轉運蛋白,可以提高微生物發酵檸檬酸能力;檸檬酸發酵培養基:玉米粉200g/l、cacl21g/l、澱粉酶20000u/l。培養基液化過程:玉米粉與水以1:4的比例混合,加入cacl2混勻。調節ph至6-6.5。加入耐高溫澱粉酶,加熱到70度保溫1h。迅速加熱到90度,保溫45-60min。玉米粉與上清液分層,加熱煮沸5-10min,用6層紗布過濾去渣滓。用生理鹽水洗下孢子,並用血球計數板計數,接種至培養基中,終濃度為5×104/ml。每隔24小時檢測糖消耗情況和檸檬酸產量。還原糖採用sdg-iv型全自動還原糖測定儀進行測定,總糖可以先用酸水解成還原糖後再測定。總糖的處理方法:取1ml發酵液上清,向其中加入6m鹽酸5ml,蒸餾水4ml,在沸水中加熱30分鐘,取1-2滴置於白瓷盤上,加1滴i-ki溶液檢測是否完全水解。如已經水解完全,則不呈現藍色。水解完畢後,冷卻至室溫,加入酚酞作為指示劑,用6m氫氧化鈉中和至微紅色,最後定容至20ml。有機酸的測定方法:採用高效液相色譜法,發酵液上清稀釋10倍,煮沸變性後,高速離心10分鐘,取上清。檢測條件:柱溫為35度,流動相5mm硫酸,流速0.5ml/min,進樣量20μl。將粗糙脈孢菌hgt1/hgt-2在黑麴黴和嗜熱毀絲黴中表達,可以提高檸檬酸發酵能力。黑麴黴工程菌比出發菌檸檬酸發酵水平提高了30%以上,糖利用加快了20%以上;嗜熱毀絲黴工程菌檸檬酸發酵水平和糖利用速率相比出發菌都提高了20%以上;在本發明提及的所有文獻都在本申請中引用作為參考,就如同每一篇文獻被單獨引用作為參考那樣。此外應理解,在閱讀了本發明的上述講授內容之後,本領域技術人員可以對本發明作各種改動或修改,這些等價形式同樣落於本申請所附權利要求書所限定的範圍。序列表中國科學院天津工業生物技術研究所一種提高絲狀真菌木質纖維素酶系表達及生物基化學品生產的方法p2016-032523patentinversion3.511662dna粗糙脈胞孢黴(neurosporacrassa)misc_feature(1)..(1662)hgt-11atgtcgtccgcccacgagaaggagacggctcctacagcagcccacattgggctcgctcac60acccaagatgtcgaacgcatcgaagcccccgtcacatggaaggcctacctcatttgcgcc120ttcgcctcattcggtggtatcttcttcggttatgactccggttatatcaacggcgttctc180ggctcgcagatcttcatcgatgccgtcgaaggcacctcccccgtccgcgatgcgctttct240gaatcccaccagtccctcgtcgtctctatcctgtcctgcggtaccttctttggcgccctc300attgccggtgatctcgccgacatgatcggccgcaagtggaccgtcattctcggctgtctc360atctacctgattggttgcgtgatccagatgatcactggtcttggtgatgctctcggtgct420atcgttgctggccgtcttattgctggtatcggtgtgggtttcgagtccgccgtcgtcatt480ctgtacatgtctgagatttgccctcgcaaggtccgtggtgccttggtcgccggttaccaa540ttctgcatcaccatcggtcttatgcttgcttcttgcgtcgtctacgcaactcaggacagg600ccggataccggtgcttaccgtattcctatcgccatccagttcatctgggctctcatcctt660gccggtggtctcatgtgtcttcccgactctccccggtacttcgtcaagaagggtaacctc720gctgctgctaccagctctctgtcccgtcttcgtggccaggaccccaactccgagtacatc780caggtcgagcttgctgagatcgtcgccaacgaggagtacgagcgtcagcttattccctcc840accacctggttcggctcctgggccaactgcttcaagggttctctctggaaggccaactcc900aacctccgcaagactattcttggtacctctcttcagatgatgcagcaatggaccggtgtc960aacttcatcttctactactccacccctttccttaagtctaccggtgccatctccaacacc1020ttcttgatctccatggtcttcaccatcatcaacgtcttctccactcccatctccttctgg1080acggtcgagcgcttcggccgccgcactatcctcttctggggtgccctcggcatgctcatc1140tgccagttcctcgtcgccattatcggtgtcaccgtcggcttcaacaagacccacatgggc1200cccgacggcgagtccatggccaacaatgtcagcgccgtaaacgcccagattgccttcatc1260gccatctttatcttcttcttcgcctccacctggggtcctggtgcttggatcttgattggc1320gagatcttccctcttcccatccgttcgcgcggtgttggtctgtccactgcctccaactgg1380ctctggaacaccatcatcgccgtcatcaccccttacatggttggcgagcagcgcggcaac1440cttaaatcgtccgtcttcttcgtctggggcggtctttgcacctgtgccttcatctacact1500tacttcctcgtgcctgagaccaagggactctcgctcgagcaggtcgacaagatgatggag1560gagacgacccccaggacttcggccaagtggaagcctcgcaccacctttgcccacgcggct1620gaggatggtatgctcaacgtacctgctgagaagcaggtttaa16622553prt粗糙脈胞孢黴(neurosporacrassa)peptide(1)..(553)hgt-12metserseralahisglulysgluthralaprothralaalahisile151015glyleualahisthrglnaspvalgluargileglualaprovalthr202530trplysalatyrleuilecysalaphealaserpheglyglyilephe354045pheglytyraspserglytyrileasnglyvalleuglyserglnile505560pheileaspalavalgluglythrserprovalargaspalaleuser65707580gluserhisglnserleuvalvalserileleusercysglythrphe859095pheglyalaleuilealaglyaspleualaaspmetileglyarglys100105110trpthrvalileleuglycysleuiletyrleuileglycysvalile115120125glnmetilethrglyleuglyaspalaleuglyalailevalalagly130135140argleuilealaglyileglyvalglyphegluseralavalvalile145150155160leutyrmetsergluilecysproarglysvalargglyalaleuval165170175alaglytyrglnphecysilethrileglyleumetleualasercys180185190valvaltyralathrglnaspargproaspthrglyalatyrargile195200205proilealaileglnpheiletrpalaleuileleualaglyglyleu210215220metcysleuproaspserproargtyrphevallyslysglyasnleu225230235240alaalaalathrserserleuserargleuargglyglnaspproasn245250255serglutyrileglnvalgluleualagluilevalalaasngluglu260265270tyrgluargglnleuileproserthrthrtrppheglysertrpala275280285asncysphelysglyserleutrplysalaasnserasnleuarglys290295300thrileleuglythrserleuglnmetmetglnglntrpthrglyval305310315320asnpheilephetyrtyrserthrpropheleulysserthrglyala325330335ileserasnthrpheleuilesermetvalphethrileileasnval340345350pheserthrproileserphetrpthrvalgluargpheglyargarg355360365thrileleuphetrpglyalaleuglymetleuilecysglnpheleu370375380valalaileileglyvalthrvalglypheasnlysthrhismetgly385390395400proaspglyglusermetalaasnasnvalseralavalasnalagln405410415ilealapheilealailepheilephephephealaserthrtrpgly420425430proglyalatrpileleuileglygluilepheproleuproilearg435440445serargglyvalglyleuserthralaserasntrpleutrpasnthr450455460ileilealavalilethrprotyrmetvalglygluglnargglyasn465470475480leulysserservalphephevaltrpglyglyleucysthrcysala485490495pheiletyrthrtyrpheleuvalprogluthrlysglyleuserleu500505510gluglnvalasplysmetmetglugluthrthrproargthrserala515520525lystrplysproargthrthrphealahisalaalagluaspglymet530535540leuasnvalproalaglulysglnval54555031584dna粗糙脈胞孢黴(neurosporacrassa)misc_feature(1)..(1584)hgt-23atgggcttgtcactcaagaagcctgaaggtgtgccgggcaagtcatggcccgccattgtc60attggcttgtttgtcgcctttggtggtgtactctttgggtatgacactggcactattggc120ggtatccttgctatgccctattggcaagatttgttttcgacaggttacagaaacccagag180catcacttggacgttaccgcgtcgcagtctgccactatcgtctccattctgtctgctgga240accttctttggcgctcttggtgccgctccccttgccgactgggctggacgacgcttgggc300cttattctgtcgtcgtttgtgtttatcttcggtgtcatcctgcagaccgcagccgtcagc360attcctctttttctggctggccgattctttgctggattgggagttggtctcatatcggca420accatccccctctatcaatccgagactgccccgaaatggattcgtggtgtcatcgtcggg480tcctatcagctagccattaccatcggtcttcttcttgcctccattgtcaacaatgccacg540cataacatgcagaacaccggctgctatcgcatccccatagctgtccaatttgcatgggcg600atcatcctgatcgttggcatgatcattcttcccgaaactccacgctttcatatcaagaga660gacaatctcccagccgccactaggtctctagctatcctccgccgtctggagcagaaccat720ccagcgatcatcgaagagctttccgagatccaagccaatcatgaatttgagaagagcctc780gggaaggcgacctacttggactgcctcaagggcaatttactcaagcggctccttactggc840tgttttctccagagcctgcagcagttgactggcatcaactttatcttctactacggcaca900cagttcttcaaaaactccggattctcagactcgtttctgatatccttgatcactaatctt960gtcaatgtcgtgtcgacccttcccggactctacgccatcgacaaatggggccggaggcct1020gttttactctggggagctgttgggatgtgtgtctgccagttcatcgttgctattcttggg1080acaacaacgacaagtcaagatgcaagcggaatgatcattgtgcataatctcgccgcacag1140aaagcagctattgcattcatctgcttctacatctttttcttcgctgcatcttggggtcca1200gttgcctgggtcgttacaggcgagatcttcccccttaaagtccgcgccaagtcgctctcc1260ataactacagcgtcgaattggctgctcaactgggccattgcttacagcacaccttacctt1320gtcaactacggccctggcaatgcgaacctgcagtccaagatcttcttcgtctggggcgga1380tgctgcttcatctgcatcgcattcgtttacttcatgatctatgagacaaaaggtctcaca1440ctggagcaggttgacgagctatatgaagaggtctcggatgccaggaagagtattggttgg1500gtgccgaccatcactttccgggagatccgggaggaaaagaaagtaagggatccagttgtt1560gatatcactgaagaggcagcttga15844527prt粗糙脈胞孢黴(neurosporacrassa)peptide(1)..(527)hgt-24metglyleuserleulyslysprogluglyvalproglylyssertrp151015proalailevalileglyleuphevalalapheglyglyvalleuphe202530glytyraspthrglythrileglyglyileleualametprotyrtrp354045glnaspleupheserthrglytyrargasnprogluhishisleuasp505560valthralaserglnseralathrilevalserileleuseralagly65707580thrphepheglyalaleuglyalaalaproleualaasptrpalagly859095argargleuglyleuileleuserserphevalpheilepheglyval100105110ileleuglnthralaalavalserileproleupheleualaglyarg115120125phephealaglyleuglyvalglyleuileseralathrileproleu130135140tyrglnsergluthralaprolystrpileargglyvalilevalgly145150155160sertyrglnleualailethrileglyleuleuleualaserileval165170175asnasnalathrhisasnmetglnasnthrglycystyrargilepro180185190ilealavalglnphealatrpalaileileleuilevalglymetile195200205ileleuprogluthrproargphehisilelysargaspasnleupro210215220alaalathrargserleualaileleuargargleugluglnasnhis225230235240proalaileileglugluleusergluileglnalaasnhisgluphe245250255glulysserleuglylysalathrtyrleuaspcysleulysglyasn260265270leuleulysargleuleuthrglycyspheleuglnserleuglngln275280285leuthrglyileasnpheilephetyrtyrglythrglnphephelys290295300asnserglypheseraspserpheleuileserleuilethrasnleu305310315320valasnvalvalserthrleuproglyleutyralaileasplystrp325330335glyargargprovalleuleutrpglyalavalglymetcysvalcys340345350glnpheilevalalaileleuglythrthrthrthrserglnaspala355360365serglymetileilevalhisasnleualaalaglnlysalaalaile370375380alapheilecysphetyrilephephephealaalasertrpglypro385390395400valalatrpvalvalthrglygluilepheproleulysvalargala405410415lysserleuserilethrthralaserasntrpleuleuasntrpala420425430ilealatyrserthrprotyrleuvalasntyrglyproglyasnala435440445asnleuglnserlysilephephevaltrpglyglycyscyspheile450455460cysilealaphevaltyrphemetiletyrgluthrlysglyleuthr465470475480leugluglnvalaspgluleutyrglugluvalseraspalaarglys485490495serileglytrpvalprothrilethrphearggluilearggluglu500505510lyslysvalargaspprovalvalaspilethrgluglualaala515520525549dna寡核苷酸5gtaacgccagggttttcccagtcacgacgcccaccccagacccagcact49656dna寡核苷酸6atccacttaacgttactgaaatctccaacacatggtgtttgttgtgactcttgtgt56754dna寡核苷酸7ctccttcaatatcatcttctgtctccgacagctgcatattttgagctcctcgtg54849dna寡核苷酸8gcggataacaatttcacacaggaaacagccgcacacgtccgcacttcct49930dna寡核苷酸9gtcggagacagaagatgatattgaaggagc301029dna寡核苷酸10gttggagatttcagtaacgttaagtggat291120dna寡核苷酸11cgcacacgtccgcacttcct201218dna寡核苷酸12tgcaataggtcaggctct181349dna寡核苷酸13gtaacgccagggttttcccagtcacgacgtagctacctacctcgagacg491449dna寡核苷酸14atccacttaacgttactgaaatctccaactgtagtacaggaggctttgc491554dna寡核苷酸15ctccttcaatatcatcttctgtctccgacagctgcccatctaaaggaacagtcg541649dna寡核苷酸16gcggataacaatttcacacaggaaacagcgacacagttcgaatagcagc491720dna寡核苷酸17gacacagttcgaatagcagc201827dna寡核苷酸18cggactagtatgtcgtccgcccacgag271926dna寡核苷酸19ccggaattcaacctgcttctcagcag262029dna寡核苷酸20ggactagtatgggcttgtcactcaagaag292130dna寡核苷酸21ccggaattcagctgcctcttcagtgatatc302230dna寡核苷酸22cggactagtatgggcttgtcactcaagaag302328dna寡核苷酸23cgcggatcctcaagctgcctcttcagtg28當前第1頁12