一種植物葉片最大生理電容和最大緊張度的測定方法與流程
2023-06-15 01:20:46 6
本發明屬於植物生理信息檢測技術領域,具體涉及一種植物葉片最大生理電容和最大緊張度的測定方法。
背景技術:
目前植物電生理信息檢測方面取得了長足的進展,人們正試圖利用植物組織電參數值的變化反映植物體宏觀與微觀的結構變化,表徵植物生命活動過程。植物屬於生物體,是介於導體和絕緣體之間的電介質,在外電場的作用下具有特定的電容值。水是植物生命周期活動中各項生理活動和酶促反應進行必不可少的成分。植物的各項生命活動離不開水的參與。當植物葉片細胞失水如蒸騰時,葉肉細胞的細胞壁、細胞都因失水而收縮,細胞體積變小。如果植物吸收水分,外液中的水分就會進入葉肉細胞,細胞因吸水而膨脹,細胞體積變大。細胞的水分狀況與細胞的這種膨脹度或收縮度緊密相關。我們把因水分的變化而導致細胞的這種膨脹度或收縮度的變化稱為葉片緊張度的變化,也即葉片細胞的這種膨脹度或收縮度稱為葉片緊張度。
細胞液濃度以及體積的變化能夠用生理電容或者緊張度來反映。植物葉片最大生理電容和最大緊張度分別表徵細胞液溶質濃度為0時的植物葉片生理電容和葉片緊張度,這種狀況因在實踐中不存在,因此,不能用儀器直接測得。本發明則是提供一種利用不同水平的鹽逆境造成水分狀況的差異,構建模型,計算出細胞液溶質濃度為0時的植物葉片生理電容和葉片緊張度。植物葉片最大生理電容和最大緊張度是每種植物固有的特性,是用植物葉片生理電容或者葉片緊張度研究植物細胞的水分變化必不可少的參照,對用植物電生理信息研究植物抗鹽、抗旱和抗逆性等抗性能力方面具有重要的意義。
技術實現要素:
本發明提供了一種測定植物葉片最大電容和最大緊張度的方法,解決了因細胞液溶質濃度為0的狀態在實際情況下不存在,難以直接用儀器測得植物葉片最大生理電容和葉片最大緊張度的問題,為用植物葉片生理電容或者葉片緊張度研究植物細胞的水分變化提供了參照。
本發明採取以下技術方案:
一種植物葉片最大生理電容和最大緊張度的測定方法,包括以下步驟:
步驟一,實驗室內採用同樣規格的穴盤萌發植物種子,配製培養液培養幼苗至3葉期以上,選擇生長較為一致的幼苗作為被考察植物幼苗;
步驟二,將被考察植物幼苗分別培養在含有不同鹽度水平的培養液中;
步驟三,待被考察植物培養至二周以上,以第一展開葉為考察對象,於同一時段測定其葉水勢w和生理電容cp;
步驟四,依據葉水勢w和生理電容cp計算葉片緊張度lt;
步驟五,利用兩參數的指數衰減方程構建葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型;
步驟六,依據葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型獲取植物葉片最大生理電容cpmax和植物葉片的最大緊張度ltmax。
進一步,所述步驟二中鹽度水平以培養液的電導率表示。
進一步,所述步驟四中計算葉片緊張度lt的公式為其中i系解離係數,r為氣體常數,t為熱力學溫度,ε0為真空介電常數,a為細胞液溶質的相對介電常數,m為細胞液溶質的相對分子質量。
進一步,所述步驟五中葉片生理電容cp與鹽逆境水平模型為葉片緊張度lt與鹽逆境水平模型為其中a1、a2、b1、b2是模型常數,xec為鹽逆境水平。
進一步,所述步驟六具體為:將xec=0分別代入和中,求出cp和lt的值,此時cp=a1,lt=a2;即a1為植物葉片最大生理電容cpmax,a2為植物葉片的最大緊張度ltmax。
本發明的有益效果為:
1)本發明建立了植物不同鹽水平下,葉片生理電容與鹽度水平以及葉片緊張度與鹽度水平模型,可以在測定植物葉片最大生理電容和植物葉片的最大緊張度的同時,描述植物電生理指標與鹽度之間的關係。
2)本發明實現了植物葉片最大生理電容和植物葉片的最大緊張度的測定,測得的植物葉片最大電容和最大緊張度是特定植物葉片生理電容或者葉片緊張度的參照值;本發明採用物理方法,對植株損傷小,結果靈敏、精確度高,可以真實反映植物活體的生理電容和緊張度。
附圖說明
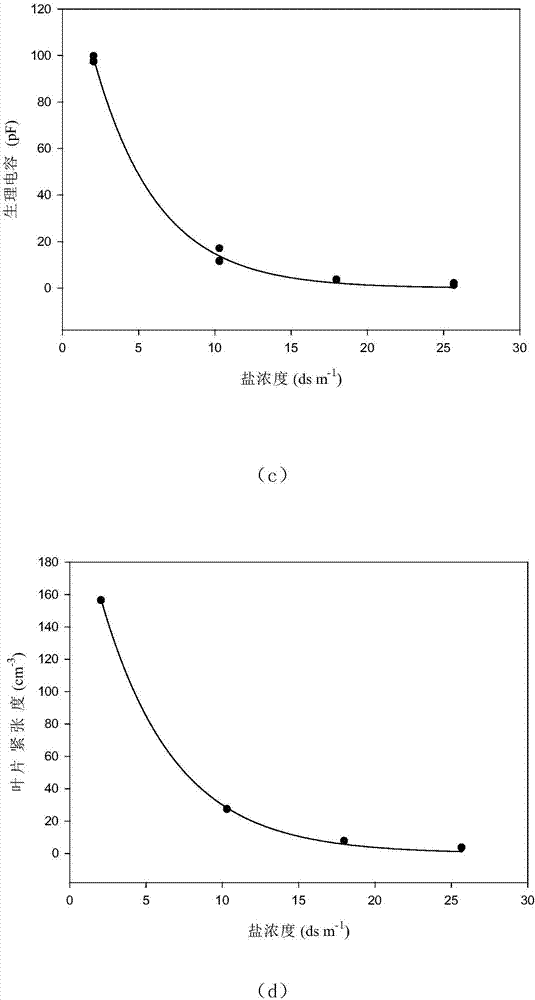
圖1為葉片生理電容cp、葉片緊張度lt與鹽逆境水平之間關係的擬合曲線圖,圖1(a)為紅秋葵葉片生理電容cp與鹽逆境水平之間關係的擬合曲線圖,圖1(b)為紅秋葵葉片緊張度lt與鹽逆境水平之間關係的擬合曲線圖,圖1(c)為綠秋葵葉片生理電容cp與鹽逆境水平之間關係的擬合曲線圖圖1(d)為綠秋葵葉片緊張度lt與鹽逆境水平之間關係的擬合曲線圖;
具體實施方式
下面將對本發明作進一步的說明,但本發明的保護範圍並不限於此。
發明原理:
植物出現乾旱時往往導致組織內體液濃度升高,電阻增大和電容值減小的現象。但是,由於植物葉片結構和內部溶液分布比較複雜,因此,乾旱引起的葉片生理特性與電物理特徵並不存在簡單的線性關係。以葉片為平板電容器的介質,葉片的水分發生變化必然引起介電常數的變化,從而在電容值上反映出來。
植物體內各項生命活動的進行離不開水,如礦質元素的運輸、吸收,光合產物的合成、轉化和運輸,植物體正常體溫維持等方面具有重要作用。植物葉片是植物進行光合作用和蒸騰作用的重要場所。葉肉細胞擴張和葉片生長對水分條件十分敏感,植物葉片要保持挺立狀態既需要纖維素的支持,還需要組織內較高膨壓的支持,植物缺水時出現的萎蔫現象就是膨壓下降的表現。當正常植株受鹽分脅迫時,葉片葉肉細胞的細胞和細胞壁因失水而收縮,細胞體積變小,且細胞內的這種水分狀況與細胞膨脹度或收縮度密切相關。葉片細胞發生的這種細胞膨脹度或收縮度可以用葉片緊張度來表示。
植物對鹽脅迫會產生應激反應,不同濃度梯度的鹽脅迫使植株較正常植株在葉片生理特徵和電物理特徵上表現出差異。實驗發現,採用電導率表徵鹽脅迫程度,葉片水勢w和生理電容cp之間存在一定的相關性。依據水勢w和生理電容cp可以用來計算出葉片緊張度lt。
兩參數的指數衰減方程為:
y=ae-bx(1)
其中,e是自然對數常數,a代表初始葉片緊張度或者生理電容,b代表衰減速率。
利用兩參數的指數衰減方程分別構建葉片生理電容cp、葉片緊張度lt與鹽度水平模型:
其中a1、a2、b1、b2是方程的常數,xec為鹽度水平,以溶液的電導率表示。
當細胞內是純水時,細胞的電容值和緊張度值最大。因此將xec=0分別代入和中,求出cp和lt的值,即分別為植物葉片最大生理電容cpmax和植物葉片的最大緊張度ltmax。
實施例:
取秋葵為研究材料,品種分別為紅秋葵和綠秋葵;實驗室內採用12孔穴盤萌發秋葵種子,配製霍格蘭培養液培養幼苗至3葉期後,分別選擇生長較為一致的幼苗作為被考察植物幼苗;添加等量的nacl和cacl2到霍格蘭培養液中,配製不同鹽度水平的培養液(用培養液的電導率表示),如表1。
表1不同鹽度水平
分別用不同鹽度水平的培養液對上述3葉期的被考察植物幼苗同時進行培養,每天更換新的相對應的培養液,以第一展開葉為考察對象,在培養的第21天的上午9:00-11:00時測定其葉水勢w和生理電容cp(表2);依據葉水勢w和生理電容cp計算葉片緊張度lt,計算葉片緊張度lt的公式為:
其中:w為植物組織水勢,mpa;i系解離係數,其值為1;r為氣體常數,0.0083l·mpa/mol·k;t為熱力學溫度k,t=273+t℃,t為環境溫度;cp為植物葉片生理電容,f;真空介電常數ε0=8.854×10-12f/m;a為細胞液溶質的相對介電常數,f/m;m為細胞液溶質的相對分子質量;葉片細胞液溶質假定為蔗糖,此時a為3.3f/m,m為342;結果如表2。
表2不同鹽度水平下兩種秋葵葉片生理電容cp、葉水勢w和葉片緊張度lt
隨後,利用兩參數的指數衰減方程構建葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型,即式(2)、(3);
通過sigmaplot軟體對葉片生理電容cp、葉片緊張度lt與鹽逆境水平之間的關係進行曲線擬合,擬合曲線見圖1;同時可得到兩參數的指數衰減方程的擬合參數,即紅秋葵為a1=123.837,b1=0.142,a2=200.706,b2=0.131;綠秋葵為a1=161.075,b1=0.239,a2=240.708,b2=0.208;將參數值代入指數衰減方程即可得到對應的關係模型;結果如表3。
表3兩種秋葵葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型
從表3可以看出,葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型都可以很好地表徵電生理指標與鹽度之間的關係,從決定係數的平方值(r2)來看,與葉片生理電容cp與鹽度關係相比,葉片緊張度lt與鹽度關係可更好地用兩參數的指數衰減方程來擬合。
依據葉片生理電容cp與鹽度水平以及葉片緊張度lt與鹽度水平模型獲取植物葉片最大生理電容cpmax和植物葉片的最大緊張度ltmax;即將xec=0分別代入式(2)和式(3)中,求出cp和lt的值,此時cp=a1,lt=a2;即a1為植物葉片最大生理電容cpmax,a2為植物葉片的最大緊張度ltmax;結果如表4所示。
表4兩種秋葵葉片最大生理電容cpmax和葉片最大緊張度ltmax
從表4可以看出,綠秋葵的最大生理電容與最大緊張度都大於紅秋葵,這與綠秋葵的葉細胞大於紅秋葵葉細胞有關;葉細胞越小抗旱抗鹽能力越強,這與紅秋葵抗逆能力強有關。
以上說明僅僅為本發明的較佳實施例,本發明並不限於列舉上述實施例,應當說明的是,任何熟悉本領域的技術人員在本說明書的教導下,所做出的所有等同替代、明顯變形形式,均落在本說明書的實質範圍之內,理應受到本發明保護。