適於酵母中的異源基因表達的啟動子的製作方法
2023-10-05 15:06:29 3
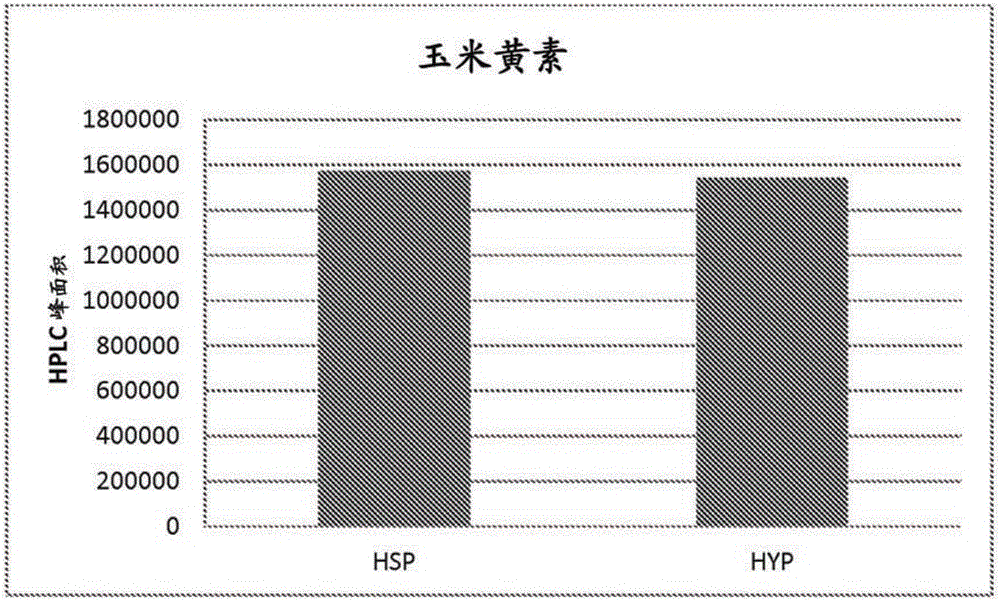
本申請要求申請日為2014年1月31日的美國臨時專利申請No.61/933,979的申請日權益,其公開內容通過提述在此併入本文。發明領域本發明涉及新的啟動子用於異源基因表達的用途,優選為用於耶氏酵母屬(genusYarrowia)的生物中基因表達的用途,涉及經遺傳修飾的耶氏酵母屬生物,並涉及用於通過培養所述經遺傳修飾的生物來生產生物合成產物的方法。發明背景細胞中天然代謝過程產生多種生物合成產物,例如精細化學品,例如尤其是胺基酸、維生素、類胡蘿蔔素,其用於多種工業分支中,包括人與動物食品、化妝品、以及製藥工業。其大規模生產是部分通過生物技術方法用微生物來進行的,所述微生物經過開發以產生和分泌大量具體所需的物質。例如,類胡蘿蔔素是在細菌、藻類和真菌中從頭合成的。近年來,越來越多地嘗試利用油質(oleaginous)酵母和真菌作為生產精細化學品,尤其是生產維生素和類胡蘿蔔素的生物。類胡蘿蔔素(如葉黃素、玉米黃素、蝦青素和β胡蘿蔔素)作為所謂的油性樹脂提取自例如耶氏酵母屬。這些油性樹脂既用作膳食補充劑的成分,也用於飼料行業(sector)中。酮式類胡蘿蔔素(ketocarotenoids),意即包含至少一個酮基的類胡蘿蔔素,例如蝦青素、角黃素、海膽酮、3-羥基海膽酮、3』-羥基海膽酮、金盞花紅素(adonirubin)和金盞花黃素(adonixanthin),是天然抗氧化劑和色素,其由一些藻類、生物和微生物作為次級代謝物產生。這些分子在能生產其的生物(如酵母和細菌)中的生物合成已得到了詳細表徵(Weinheim等,(1996)Ullmann'sEncyclopediaofIndustrialChemistry, "Vitamins",Vol.A27,pp.443-613;BiochemicalPathways:AnAtlasofBiochemistryandMolecularBiology,JohnWiley&Sons,Michal,G.Ed.(1999);Ong,A.S.,Niki,E.和Packer,L.,(1995)"Nutrition,Lipids,HealthandDisease"ProceedingsoftheUNESCO/ConfederationofScientificandTechnologicalAssociationsinMalaysiaandtheSocietyforRreeRadicalResearch-Asia,舉辦於1994年9月1-3日,於Penang,Malaysia,AOCSPress,Champaign,ILX,374S)。具體而言,類胡蘿蔔素在耶氏酵母中的生物合成描述於WO2006/102342中。由於酮式類胡蘿蔔素的著色特性,將其與特別是蝦青素用作家畜營養中(尤其是在鱒魚、鮭魚和蝦的養殖中)的著色劑。因此,用於生產天然生物合成產物(並且尤其是類胡蘿蔔素)的經濟的生物技術方法是非常重要的。WO2008/042338公開了多個用於在耶氏酵母中過表達類胡蘿蔔素生物合成基因的啟動子。EP0220864A中公開了一種的啟動子。該文獻描述了解脂耶氏酵母(Yarrowialipolytica)啟動子XPR2。該XPR2酵母啟動子在缺乏偏好的碳和氮源的培養基上僅在pH6.0以上有活性,並且完全誘導需要在培養基中高水平的蛋白腖(Ogrydziak,D.M.,Demain,A.L.,和Tannenbaum,S.R.,(1977)Biochim.Biophys.Acta.497:525-538.;Ogrydziak,D.M.和Scharf,S.J.,(1982)Gen.Microbiol.128:1225-1234.)。用於在酵母(例如耶氏酵母)中表達基因的另一種啟動子,描述於WO1997/044470中。然而,迄今為止使用的啟動子無法完全滿足在耶氏酵母中進行高表達的需求。因此需要提供更好地滿足這些需求的啟動子。因此,本發明的目的是提供新的改進的酵母啟動子,尤其用於在酵母中克隆表達,而且用於在選擇的表達系統中異源表達上述合意的精細化學品。發明概述本發明涉及一種重組核酸分子,其包含:a)與SEQIDNO:1(在此稱為HSP)或SEQIDNO:2(在此稱為HYP)的多核苷酸序列具有至少90%同一性的多核苷酸序列,或b)在嚴格條件下與核酸序列SEQIDNO:1或SEQIDNO:2雜交的核酸序列,或c)a)或b)的序列的功能上等同的片段。在一個方面,本發明涉及一種重組核酸分子,其包含與SEQIDNO:1或SEQIDNO:2的多核苷酸序列具有至少95%序列同一性的多核苷酸序列。在本發明的一個具體方面,所述重組核酸分子作為啟動子發揮功能。所述重組核酸分子用於在酵母中調控基因的表達。在一個實施方案中,所述酵母是耶氏酵母屬的生物。在一個實施方案中,所述酵母是解脂耶氏酵母的菌株。在另一個實施方案中,所述被調控的基因對於耶氏酵母屬的生物體是異源的。在一個實施方案中,所述重組核酸分子與編碼蛋白的基因是功能上相連的(functionallylinked)。在一個實施方案中,所述基因與一個或多個其他調控信號是功能上相連的。在一個具體實施方案中,所述功能上相連的基因選自編碼蛋白的核酸的組,所述蛋白來自:a)有機酸的生物合成途徑,或b)脂質和脂肪酸的生物合成途徑,或c)二醇的生物合成途徑,或d)芳香族化合物的生物合成途徑,或e)維生素的生物合成途徑,或f)類胡蘿蔔素,特別是酮式類胡蘿蔔素的生物合成途徑。在一個實施方案中,本發明所述的重組核酸分子是載體。本發明還涉及經遺傳修飾的微生物,其包含上述的重組核酸分子。在一個實施方案中,所述微生物是耶氏酵母屬的成員。在另一個實施方案中,上述的重組核酸分子用於在宿主微生物中表達一個或多個基因,其中至少一個或多個基因的表達速率(expressionrate)與野生型相比是提高的。在一個具體實施方案中,所述基因在宿主微生物中的表達調控通過如下實現:a)將一個或多個上述重組核酸分子引入宿主微生物的基因組中,從而使宿主微生物一個或多個內源基因的表達在引入的重組核酸分子控制下發生;b)將一個或多個基因引入所述微生物的基因組中,從而使所述一個或多個引入的基因的表達在上述重組核酸分子控制下發生,所述重組核酸分子對於宿主微生物是內源的;或c)將一個或多個核酸構建體引入宿主微生物中,其中所述一個或多個核酸構建體包含至少一個上述重組核酸分子,且所述重組核酸分子與一個或多個要表達的基因是功能上相連的。本發明還涉及生產生物合成產物的方法,包括如下步驟:a)在培養基中培養耶氏酵母屬的經遺傳修飾的微生物,其中所述經遺傳修飾的微生物包含上述的重組核酸分子,其中所述重組核酸分子與編碼蛋白的基因是功能上相連的;及b)生產步驟a)中製成的生物合成產物。在一些實施方案中,所述生物合成產物分泌至培養基中。在一些實施方案中,所述生物合成產物在微生 物中積累。在一些實施方案中,將所述生物合成產物從分離的微生物生物質回收。在一些實施方案中,所述生物合成產物是類胡蘿蔔素。附圖簡述圖1和圖2分別顯示了搖瓶實驗中測試的兩個啟動子的平均玉米黃素和蝦青素水平,以HPLC峰面積表示(對於玉米黃素:10個具有HSP啟動子的轉化體取平均值,以及5個具有HYP啟動子的轉化體取平均值。對於蝦青素生產:12個具有HSP啟動子的轉化體取平均值,以及12個具有HYP啟動子的轉化體取平均值)。圖3顯示了在發酵罐中測試的來自兩個啟動子的crtZ表達的水平。在分析的不同發酵時間點處(y軸)的所述水平以產生的蝦青素的量來描繪,以HPLC峰面積表示。圖4顯示了相比於強組成型表達的內源基因,發酵罐中表達的crtZmRNA的水平。圖5顯示了由攜帶lacZ基因的6個不同轉化體分別在HSP和HYP啟動子控制下產生的β-半乳糖苷酶單位。序列表所附的序列表中列出的核酸序列以37C.F.R.§1.822中定義的核苷酸鹼基的標準字母縮寫表示。每個核酸序列僅顯示一條鏈,但通過對所展示的鏈的任意提述,視為已包括了其互補鏈。在所附的序列表中:SEQIDNO:1顯示了來自解脂耶氏酵母的HSP啟動子:SEQIDNO:2顯示了來自解脂耶氏酵母的HYP啟動子:發明詳述本發明涉及新的重組核酸分子,其為啟動子。本發明的啟動子是指導相關編碼區(基因)轉錄的DNA區。根據本發明,所述重組核酸分子包含a)與SEQIDNO:1或SEQIDNO:2的多核苷酸序列具有至少90%同一性的多核苷酸序列,或b)在嚴格條件下與 核酸序列SEQIDNO:1或SEQIDNO:2雜交的核酸序列,或c)a)或b)的序列的功能上等同的片段。尤其優選的重組核酸分子與SEQIDNO:1或2各自的核酸序列具有至少70%,優選至少80%,至少85%,至少90%,至少92%,至少95%,至少96%,至少97%,至少98%,尤其優選至少99%的同一性。可以在SEQIDNO:1或SEQIDNO:2的核酸序列的至少100,至少200,至少300,至少400,至少500,至少800個核苷酸,或其完整序列找到百分比同一性。在一些實施方案中,上述SEQIDNO:1或SEQIDNO:2的同源物(homologue)通過核苷酸取代、插入或缺失而源自SEQIDNO:1或SEQIDNO:2的多核苷酸序列。本發明還涉及包含核酸序列的重組核酸分子,所述核酸序列在嚴格條件下與SEQIDNO:1或SEQIDNO:2的核酸序列雜交。在一個方面,本發明涉及如SEQIDNO:1或SEQIDNO:2所示的重組核酸分子,或在嚴格條件下與核酸序列SEQIDNO:1或SEQIDNO:2雜交的核酸序列,或上述序列的功能上等同的片段,其中重組核酸分子作為啟動子發揮功能。核酸序列SEQIDNO:1和SEQIDNO:2各代表克隆自解脂耶氏酵母的啟動子序列。在一些實施方案中,啟動子能用於在酵母中調控基因的表達,所述酵母包括但不限於耶氏酵母。在一個具體實施方案中,所述啟動子能用於在解脂耶氏酵母的菌株中調控基因的表達。本發明的啟動子能至少在酵母中具有啟動子活性,並且包括全長啟動子序列及其功能片段,融合序列,以及天然存在的啟動子的同源物。可以用限制性內切酶來消化本發明的核酸分子,然後進行適當的測定法以確定啟動子活性所需的最小序列。此類片段自身各自代表本發明的實施方案。啟動子的同源物與天然存在的啟動子區別在於已缺失、插入、倒位、取代和/或衍生化(derivatized)至少1、2、3個或幾個核苷酸。啟動子的同源物(至少在酵母中)能夠保留作為啟動子的活性,儘管其活性可為提高的、降低的,或依賴某種刺激物而存在的。本發明的啟動子可以包含賦予發育表達控制和調節表達控制的一個或多個序列元件。已發現,兩個新的啟動子(HSP和HYP啟動子,分別如SEQIDNO:1和SEQIDNO:2所示)及其同源物特別適於在酵母中(特別是在耶氏酵母中)使 靶基因的表達增強至相對高的水平。靶基因的例子包括生產類胡蘿蔔素或其他類異戊二烯化合物(isoprenoids)的途徑中所涉及的基因。在一些實施方案中,HSP和HYP啟動子包括其同源物能夠引起相對高濃度的類胡蘿蔔素積累。在優選的實施方案中,本發明提供兩個克隆的酵母啟動子(分別為HSP和HYP啟動子)。克隆的酵母啟動子意指通過遺傳工程中使用的標準克隆步驟克隆的酵母啟動子,從而將DNA區段從其天然位置重置至不同的位點,其會在該位點處複製。克隆過程涉及想要的DNA區段的切除和分離,將DNA片斷插入至載體分子中,以及將重組載體整合至細胞中,在此DNA區段的多個拷貝或克隆會得到複製。可以從解脂耶氏酵母菌株分離本發明的酵母啟動子DNA序列。或者,可以基於SEQIDNO:1或SEQIDNO:2中所示DNA序列所表示的DNA序列來構建本發明的啟動子序列。酵母啟動子意指結構基因的5』端處的核苷酸序列,其指導轉錄的起始。啟動子序列用於驅動下遊基因的表達。啟動子通過提供RNA聚合酶及其他起始和激活因子的結合位點來驅動轉錄。通常,啟動子優選地向下遊方向驅動轉錄。轉錄水平由啟動子調控。因此,在構建異源啟動子/結構基因組合時,將結構基因置於啟動子的調節控制之下,使得基因的表達受到啟動子序列的控制。優選地,將啟動子置於結構基因的上遊並且距離轉錄起始位點一定距離處,所述距離近似於該啟動子與其控制的基因在天然設置中的距離。如本領域已知的,可以容許此距離的一些變化而不損失啟動子功能。例如,啟動子的轉錄效率可以通過直接測量(例如通過Northern印跡或引物延伸測量)來自該啟動子的mRNA轉錄的量來確定,或是通過測量從該啟動子表達的基因產物來間接地確定。根據本發明的方法,「轉錄」意指根據該方法從DNA模板開始產生互補的RNA分子。該過程中涉及蛋白如RNA聚合酶,所謂的σ因子和轉錄調節子蛋白。然後,合成的RNA在翻譯過程中充當模板,而翻譯過程產生生物合成的活性蛋白。「功能上連接」意指在這種連結中,例如本發明的啟動子之一和要表達的核酸序列(以及如果適當的話其他調控元件(例如終止子))以這樣的方式順序安排(sequentialarrangement)使得每一個調控元件能夠在該核酸序列的表 達中達成其功能。化學意義上的直接連接對此並不是絕對必要的。遺傳控制序列例如增強子序列還能在更遠的位置或者甚至從其他DNA分子對靶序列行使其功能。這樣的排列是優選的,在該排列中要表達的核酸序列或要表達的基因被置於本發明的啟動子序列之後(即在3』端),從而使兩個序列共價連接在一起。啟動子序列與要表達的核酸序列之間的距離可以是例如少於200個鹼基對,少於100個鹼基對,或少於50個鹼基對。「表達活性」或「表達速率」意指在設定的一段時間過程中產生的蛋白的量。在本發明中,所述蛋白由基因編碼,所述基因與HSP啟動子或HYP啟動子是功能上相連的。「引發的表達活性」或「引發的表達速率」意指當蛋白是由引發而產生的時,在設定的一段時間過程中產生的該蛋白的量。在本發明中,當編碼所述蛋白的基因與與HSP啟動子或HYP啟動子是功能上相連的時,則該蛋白是由引發而產生的,當這些啟動子與所述基因在功能上不相連時,則該蛋白不以顯著量產生。生物合成的活性蛋白/酶產生的形成速率是轉錄速率和翻譯速率的乘積(product)。根據本發明可能影響兩種速率,並且因此影響微生物中產物的形成速率。根據本發明,「異源的」基因表達意指啟動子及與其功能上相連的基因在野生型生物中並非天然地以此排列存在。因此,異源性基因表達包括這樣的情形,其中啟動子、或要表達的基因、或兩種組件都不在對應的生物的野生型中天然存在;或者其中啟動子和要表達的基因二者均在野生型生物中天然存在但處於遙遠的染色體位置上,從而在野生型生物中不存在功能上的連接。根據本發明,術語「野生型」或「野生型生物」意指對應的初始生物(initialorganism)。依賴於上下文,術語「生物」可以指初始生物(野生型)或本發明經遺傳修飾的生物,例如耶氏酵母屬的生物。術語「取代」意指一個或多個核苷酸對一個或多個核苷酸的交換。「缺失」是通過直接連接來代替核苷酸。插入是將核苷酸引入至核酸序列中,其中用一個或多個核苷酸正式代替直接連接。兩個核酸之間的同一性意指在各個情況下跨越核酸全長的核苷酸的同一性,尤其是使用設置如下參數的Clustal方法(HigginsDG,SharpPM.,(1998) Fastandsensitivemultiplesequencealignmentsonamicrocomputer.ComputAppl.Biosci.5(2):151-1),藉助來自Informax(USA)的VectorNTISuite7.1軟體通過比較計算出的同一性:多重比對參數:逐對比對參數:例如,與序列SEQIDNO:1或2具有至少70%同一性的核酸序列相應地意指這樣的核酸序列,其序列與序列SEQIDNO:1或2相比(尤其是根據上述程序算法設定上述參數來比較)顯示出至少70%同一性。「雜交」意指多核苷酸或寡核苷酸在嚴格條件下與大體互補的序列結合的能力,而在這些條件下不發生與非互補對象之間的非特異性結合。對此,所述序列優選為90-100%互補的。將能夠互相特異性結合的互補序列的特性應用於例如Northern或Southern印跡技術中,或是PCR或RT-PCR的引物結合中。根據本發明,雜交在嚴格條件下發生。此類雜交條件描述於例如Sambrook,J.,Fritsch,E.F.,Maniatis,T.,in:MolecularCloning(ALaboratoryManual),第2版,ColdSpringHarborLaboratoryPress,1989,第9.31-9.57頁中,或描述於CurrentProtocolsinMolecularBiology,JohnWiley&Sons,N.Y.(1989),6.3.1-6.3.6中。嚴格雜交條件意指具體為:在溶液中42℃過夜溫育,所述溶液由50%甲 醯胺、5XSSC(750mMNaCl,75mM檸檬酸三鈉)、50mM磷酸鈉(pH7.6)、5X登哈特溶液(Denhardt'ssolution)、10%右旋糖酐硫酸酯和20g/ml變性的、剪切的(sheared)鮭魚精子DNA組成,然後用0.1XSSC在65℃洗滌濾器。「功能上等同的片段」意指對於啟動子而言,具有作為起始序列的基本上相同的啟動子活性的片段。「片段」意指本申請所述啟動子的部分的序列。尤為優選的是,使用核酸序列SEQIDNO:1或SEQIDNO:2作為啟動子,用於在耶氏酵母屬的生物中表達基因。所有前述的啟動子都可以進一步以自身已知的方式(inamannerknownperse)從核苷酸構建模塊(buildingblock)通過化學合成來產生,例如通過對雙螺旋的互補核酸構建模塊、個體重疊的片段壓縮來進行。寡核苷酸的化學合成可以按已知方式,通過亞磷醯胺(phosphoamidite)方法(Voet,Voet,(1995)Biochemistry,第2版,WileyPressNewYork,pp.896-897)來進行。用DNA聚合酶的Klenow片段及連接反應來添加合成的寡核苷酸及填補缺口,以及通用的克隆方法描述於Sambrook等(1989)Molecularcloning:Alaboratorymanual,ColdSpringHarborLaboratoryPress中。用本發明的啟動子,原則上可以在耶氏酵母屬的生物中表達任何基因。這些要在耶氏酵母屬的生物中表達的基因下文也稱為「效果基因」。優選的效果基因是來自生物合成產物的生物合成途徑的基因,所述生物合成產物可以在耶氏酵母屬的生物中產生,即在野生型中或經遺傳改造的野生型中產生。優選的生物合成產物是精細化學品。術語「精細化學品」是本領域所知的,並且包括由生物產生的並用於多種工業分支(例如但不限於製藥業、農業、化妝品業、食品及飼料業)的化合物。這些化合物包括有機酸(例如酒石酸、衣康酸、和二氨基庚二酸),脂質,飽和及不飽和脂肪酸(例如花生四烯酸),二醇(例如丙二醇和丁二醇),芳香族化合物(例如芳香胺、香草醛和靛藍(indigo)),類胡蘿蔔素和維生素及輔因子。高等動物已經失去了合成維生素、類胡蘿蔔素、輔因子和營養製品(nutraceuticals)的能力,因此需要攝入它們,儘管其他生物如細菌可容易地合成它們。這些分子或是生物活性分子本身,或是生物活性物質的前體,其充當許多代謝途徑中的中間產物或電子載體。這些化合物除了自身營養價值外 還作為著色劑、抗氧化劑和催化劑或其他加工助劑而具有重大的工業價值。術語「維生素」是本領域已知的,並且包括生物正常行使功能所需但該生物自身不能合成的營養物。維生素的組可以包括輔因子和營養製品化合物。術語「輔因子」包括正常酶活性發生所需的非蛋白化合物。這些化合物可以是有機的或無機的;本發明的輔因子優選為有機的。術語「營養製品」包括促進生物和動物(尤其是人)中健康的食品添加劑。此類分子的例子是維生素、抗氧化劑以及某些脂質(例如多不飽和脂肪酸)。優選的能夠在耶氏酵母屬的生物中生產的精細化學品或生物合成產物是類胡蘿蔔素,例如八氫番茄紅素、番茄紅素、β-胡蘿蔔素、葉黃素、玉米黃素、蝦青素、角黃素、海膽酮、3-羥基海膽酮、3』-羥基海膽酮、金盞花紅素、堇黃素(violaxanthin)和金盞花黃素。優選的類胡蘿蔔素是酮式類胡蘿蔔素,例如蝦青素、角黃素、海膽酮、3-羥基海膽酮、3』-羥基海膽酮、金盞花紅素和金盞花黃素。因此,用本發明的啟動子在酵母中表達的優選的基因選自編碼蛋白的核酸的組,所述蛋白來自:a)有機酸的生物合成途徑,b)脂質和脂肪酸的生物合成途徑,c)二醇的生物合成途徑,d)芳香族化合物的生物合成途徑,e)維生素的生物合成途徑,或f)類胡蘿蔔素,特別是酮式類胡蘿蔔素的生物合成途徑。因此,用本發明的啟動子在耶氏酵母屬生物中表達的優選的基因是編碼來自類胡蘿蔔素生物合成途徑的蛋白的基因。優選的基因選自下組:編碼酮酶的核酸、編碼[β]-羥化酶的核酸、編碼[β]-環化酶的核酸、編碼[ε]-環化酶的核酸、編碼玉米黃素環氧酶的核酸、編碼花葯黃質(antheraxanthin)環氧酶的核酸、編碼新黃素合酶的核酸、編碼HMG-CoA還原酶的核酸、編碼(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶的核酸、編碼1-脫氧-D-木糖-5-磷酸合酶的核酸、編碼1-脫氧-D-木糖-5-磷酸還原異構酶(reductoisomerase)的核酸、編碼異戊烯基-二磷酸[δ]-異構酶的核酸、編碼香葉基-二磷酸合酶的核酸、編碼法呢基-二磷酸合酶的核酸、編碼香葉基-香葉基-二磷酸合酶的核酸、編碼八氫番茄紅素合酶的核酸、編碼八氫番茄紅素去飽和酶的核酸、編碼前八氫番茄紅素(prephytoene)合酶的核酸、編碼ζ-胡蘿蔔素去飽和酶的核酸、編碼crtISO蛋白的核酸、編碼4-二磷酸胞苷-2-C-甲基-D-赤蘚醇合酶的核酸、編碼4-二磷酸胞苷-2-C-甲基-D-赤蘚醇激酶 的核酸、編碼2-甲基-D-赤蘚醇-2,4-環二磷酸合酶的核酸、和編碼羥基甲基丁烯基-二磷酸合酶的核酸。酮酶意指具有在類胡蘿蔔素的任選取代的[β]-紫羅蘭酮環上引入酮基的酶活性的蛋白。具體而言,酮酶意指具有將[β]-胡蘿蔔素轉變為角黃素的酶活性的蛋白。編碼酮酶的核酸及其對應的酮酶的例子為例如來自雨生紅球藻(Haematococcuspluvialis)、尤其是來自雨生紅球藻雨生紅球藻、橙黃土壤桿菌(Agrobacteriumaurantiacum)、產鹼菌屬(Alicaligenes)、馬氏副球菌(Paracoccusmarcusii)、集胞藻屬(Synechocystis)、慢生根瘤菌屬(Bradyrhizobium)、雨生紅球藻、副球菌屬、橙色短波單胞菌(Brevundimonasaurantiaca)、泡沫節球藻(Nodulariaspumigena)的序列。[β]-環化酶意指具有將末端線性番茄紅素殘基轉變為[β]-紫羅蘭酮環的酶活性的蛋白。具體而言,[β]-環化酶意指具有將[γ]-胡蘿蔔素轉變為[β]-胡蘿蔔素的酶活性的蛋白。[β]-環化酶基因的例子是編碼如下登錄號的[β]-環化酶的核酸:S66350番茄紅素β-環化酶,CAA60119番茄紅素合酶,AAG10429β環化酶[Yarrowiaerecta(直立耶氏酵母)],AAA81880番茄紅素環化酶,AAB53337番茄紅素β環化酶,AAL92175β-番茄紅素環化酶[橙色宮燈百合(Sandersoniaaurantiaca)],CAA67331番茄紅素環化酶[洋水仙(Narcissuspseudonarcissus)],AAM45381β環化酶[Yarrowiaerecta(直立耶氏酵母)],AAL01999番茄紅素環化酶[黃色桿菌屬菌種Py2(Xanthobactersp.Py2)],ZP_000190假設蛋白[橙色綠屈撓菌(Chloroflexusaurantiacus)],AAF78200番茄紅素環化酶[慢生根瘤菌屬菌種ORS278(Bradyrhizobiumsp.ORS278)],BAB79602crtY[成團泛菌雞血藤致病變種(Pantoeaagglomeranspv.milletiae)],CAA64855番茄紅素環化酶[灰色鏈黴菌(Streptomycesgriseus)],AAA21262番茄紅素(dycopene)環化酶[成團泛菌(Pantoeaagglomerans)],C37802crtY蛋白–噬夏孢歐文氏菌(Erwiniauredovora),BAB79602crtY[成團泛菌雞血藤致病變種],AAA64980番茄紅素環化酶[成團泛菌],AAC44851番茄紅素環化酶,BAA09593番茄紅素環化酶[副球菌屬菌種MBIC1143],CAB56061番茄紅素β-環化酶[馬氏副球菌],BAA20275番茄紅素環化酶[長赤桿菌(Erythrobacterlongus)],AAK07430番茄紅素β-環化酶[巴勒斯坦側金盞花(Adonispalaestina)],CAA67331番茄紅素環化酶[洋水仙],AAB53337番茄紅素β環化酶,BAC77673番茄紅素β-單環化酶[海洋細菌 P99-3]。羥化酶意指具有將羥基引入至類胡蘿蔔素的任選取代的[β]-紫羅蘭酮環上的酶活性的蛋白。具體而言,羥化酶意指具有將[β]-胡蘿蔔素轉變為玉米黃素或將角黃素轉變為蝦青素的酶活性的蛋白。羥化酶基因的例子是:編碼來自雨生紅球藻,登錄號AX038729WO0061764)的羥化酶;以及如下登錄號的羥化酶的核酸:CAA70427.1,CAA70888.1,CAB55625.1,AF499108-1,AF315289-1,AF296158-1,AAC49443.1,NP-194300.1,NP-200070.1,AAG10430.1,CAC06712.1,AAM88619.1,CAC95130.11AAL80006.1,AF162276-1,AA053295.1,MN85601.1,CRTZ_ERWHE,CRTZ_PANAN,BAB79605.1,CRTZ_ALCSP,CRTZ_AGRAU,CAB56060.1,ZP-00094836.1,AAC44852.1,BAC77670.1,NP-745389.1,NP-344225.1,NP-849490.1ZP-00087019.1,NP-503072.1,NP-852012.1,NP-115929.1,ZP-00013255.1。HMG-CoA還原酶意指具有將3-羥基-3-甲基戊二醯基-輔酶A轉變為甲羥戊酸的酶活性的蛋白。(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶意指具有將(E)-4-羥基-3-甲基丁-2-烯基-二磷酸轉變為異戊烯基二磷酸和二甲基烯丙基二磷酸的酶活性的蛋白。1-脫氧-D-木糖-5-磷酸合酶意指具有將羥基乙基-ThPP和甘油醛-3-磷酸轉變為1-脫氧-D-木糖-5-磷酸的酶活性的蛋白。1-脫氧-D-木糖-5-磷酸還原異構酶意指具有將1-脫氧-D-木糖-5-磷酸轉變為2-C-甲基-D-赤蘚醇-4-磷酸的酶活性的蛋白。異戊烯基-二磷酸[δ]-異構酶意指具有將異戊烯基-二磷酸轉變為二甲基烯丙基二磷酸的酶活性的蛋白。香葉基-二磷酸合酶意指具有將異戊烯基二磷酸和二甲基烯丙基二磷酸轉變為香葉基-二磷酸的酶活性的蛋白。法呢基-二磷酸合酶意指具有連續地轉變2分子的異戊烯基二磷酸及二甲基烯丙基二磷酸並將所得的香葉基-二磷酸轉變為法呢基-二磷酸的酶活性的蛋白。香葉基-香葉基-二磷酸合酶意指具有將法呢基-二磷酸轉變為香葉基-香葉基-二磷酸的酶活性的蛋白。八氫番茄紅素合酶意指具有將香葉基-香葉基-二磷酸轉變為八氫番茄紅 素的酶活性的蛋白。八氫番茄紅素去飽和酶意指具有將八氫番茄紅素轉變為六氫番茄紅素(phytofluene)和/或將六氫番茄紅素轉變為4-胡蘿蔔素(ζ-胡蘿蔔素)的酶活性的蛋白。ζ-胡蘿蔔素去飽和酶意指具有將[ζ]-胡蘿蔔素轉變為鏈孢紅素(neurosporin)和/或將鏈孢紅素轉變為番茄紅素的酶活性的蛋白。crtISO蛋白意指具有將7,9,7',9'-四-順式-番茄紅素轉變為全反式番茄紅素的酶活性的蛋白。HMG-CoA還原酶基因的例子是:編碼來自擬南芥(Arabidopsisthaliana)的HMG-CoA還原酶的核酸,其登錄號為NM-106299;以及進一步來自其他生物的具有如下登錄號的HMG-CoA還原酶基因:P54961,P54870,P54868,P54869,O02734,P22791,P54873,P54871,P23228,P13704,P54872,Q01581,P17425,P54874,P54839,P14891,P34135,O64966,P29057,P48019,P48020,P12683,P43256,Q9XEL8,P34136,O64967,P29058,P48022,Q41437,P12684,Q00583,Q9XHL5,Q41438,Q9YAS4,O76819,O28538,Q9Y7D2,P54960,051628,P48021,Q03163,P00347,P14773,Q12577,Q59468,P04035,O24594,P09610,Q58116,O26662,Q01237,O01559,Q12649,O74164,O59469,P51639,O10283,O08424,P20715,P13703,P13702,Q96UG4,Q8SQZ9,O15888,Q9TUM4,P93514,Q39628,P93081,P93080,Q944T9,Q40148,Q84MM0,Q84LS3,Q9Z9N4,Q9KLM0。(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶基因的例子是:編碼來自擬南芥(lytB/ISPH)的(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶的核酸;以及進一步來自其他生物的具有如下登錄號的(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶基因:T04781,AF270978-1,NP-485028.1,NP-442089.1,NP-681832.1,ZP-00110421.1,ZP-00071594.1,ZP-00114706.1,ISPH_SYNY3,ZP-00114087.1,ZP-00104269.1,AF398145-1,AF398146-1,AAD55762.1,AF514843-1,NP-622970.1,NP-348471.1,NP-562001.1,NP-223698.1,NP-781941.1,ZP-00080042.1,NP-859669.1,NP-214191.1,ZP-00086191.1,ISPH_VIBCH,NP-230334.1,NP-742768.1,NP-302306.1,ISPH_MYCLE,NP-602581.1,ZP-00026966.1,NP-520563.1,NP-253247.1,NP-282047.1,ZP-00038210.1,ZP-00064913.1,CAA61555.1,ZP-00125365.1,ISPH_ACICA,EAA24703.1, ZP-00013067.1,ZP-00029164.1,NP-790656.1,NP-217899.1,NP-641592.1,NP-636532.1,NP-719076.1,NP-660497.1,NP-422155.1,NP-715446.1,ZP-00090692.1,NP-759496.1,ISPH_BURPS,ZP-00129657.1,NP-215626.1,NP-335584.1,ZP-00135016.1,NP-789585.1,NP-787770.1,NP-769647.1,ZP-00043336.1,NP-242248.1,ZP-00008555.1,NP-246603.1,ZP-00030951.1,NP-670994.1,NP-404120.1,NP-540376.1,NP-733653.1,NP-697503.1,NP-840730.1,NP-274828.1,NP-796916.1,ZP-00123390.1,NP-824386.1,NP-737689.1,ZP-00021222.1,NP-757521.1,NP-390395.1,ZP-00133322.1,CAD76178.15NP-600249.1,NP-454660.1,NP-712601.1,NP-385018.1,NP-751989.1。1-脫氧-D-木糖-5-磷酸合酶基因的例子是:編碼來自番茄(Lycopersiconesculentum)的1-脫氧-D-木糖-5-磷酸合酶的核酸,以及進一步來自其他生物的具有如下登錄號的1-脫氧-D-木糖-5-磷酸合酶基因:AF143812-1,DXS_CAPAN,CAD22530.1,AF182286-1,NP-193291.1,T52289,AAC49368.1,AAP14353.1,D71420,DXS_ORYSA,AF443590-1,BAB02345.1,CAA09804.2,NP-850620.1,CAD22155.2,AAM65798.1,NP-566686.1;CAD22531.1,AAC33513.1,CAC08458.1,AAG10432.1,T08140,AAP14354.1,AF428463-1,ZP-00010537.1,NP-769291.1,AAK59424,1,NP-107784.1,NP-697464.1,NP-540415.1,NP-196699.1,NP-384986.1,ZP-00096461.1,ZP-00013656.1,NP-353769.1,BAA83576.1,ZP-00005919.1,ZP-00006273.1,NP-420871.1,AAM48660.1,DXS_RHOCA,ZP-00045608.1,ZP-00031686.1,NP-841218.1,ZP-00022174.1,ZP-00086851.1,NP-742690.1,NP-520342.1,ZP-00082120.1,NP-790545.1,ZP-00125266.1,CAC17468.1,NP-252733.1,ZP-00092466.1,NP-439591.1,NP-414954.1,NP-752465.1,NP-622918.1,NP-286162.1,NP-836085.1,NP-706308.1,ZP-00081148.1,NP-797065.1,NP-213598.1,NP-245469.1,ZP-00075029.1,NP-455016.1,NP-230536.1,NP-459417.1,NP-274863.1,NP-283402.1,NP-759318,1,NP-406652.1,DXS_SYNLE,DXS_SYNP7,NP-440409.1,ZP-00067331.1,ZP-00122853.17NP-717142.1,ZP-00104889.1,NP-243645.1,NP-681412.1,DXSSYNEL,NP-637787.1,DXS-CHLTE,ZP-00129863.1,NP-661241.1,DXS_XANCP,NP-470738.1,NP-484643.1,ZP-00108360.1,NP-833890.1,NP-846629.1,NP-658213.1, NP-642879.1,ZP-00039479.1,ZP-00060584.1,ZP-00041364.1,ZP-00117779.1,NP-299528.1。1-脫氧-D-木糖-5-磷酸還原異構酶基因的例子是:編碼來自擬南芥的1-脫氧-D-木糖-5-磷酸還原異構酶的核酸,以及進一步來自其他生物的具有如下登錄號的1-脫氧-D-木糖-5-磷酸還原異構酶基因:AF148852,AY084775,AY054682,AY050802,AY045634,AY081453,AY091405,AY098952,AJ242588,AB009053,AY202991,NP-201085.1,T52570,AF331705-1,BAB16915.1,AF367205-1,AF250235-1,CAC03581.1,CAD22156.1,AF182287-1,DXR_MENPI,ZP-00071219.1,NP-488391.1,ZP-00111307.1,DXR_SYNLE,AAP56260.1,NP-681831.1,NP-442113.1,ZP-00115071.1,ZP-00105106.1,ZP-00113484.1,NP-833540.1,NP-657789.1,NP-661031.1,DXR-BACHD,NP-833080.1,NP-845693.1,NP-562610.1,NP-623020.1,NP-810915.1,NP-243287.1,ZP-00118743.1,NP-464842.1,NP-470690.1,ZP-00082201.1,NP-781898.1,ZP-00123667.1,NP-348420.1,NP-604221.1,ZP-00053349.1,ZP-00064941.1,NP-246927.1,NP-389537.1,ZP-00102576.15NP-519531.1,AF124757-19,DXR_ZYMMO,NP-713472.1,NP-459225.1,NP-454827.1,ZP-00045738.1,NP-743754.1,DXR_PSEPK,ZP-00130352.1,NP-702530.1,NP-8417441,NP-438967.1,AF514841-1,NP-706118.1,ZP-00125845.1,NP-404661.1,NP-285867.1,NP-240064.1,NP-414715.1,ZP-00094058.1,NP-791365.1,ZP-00012448.1,ZP-00015132.1,ZP-00091545.1,NP-629822.1,NP-771495.1,NP-798691.1,NP-231885.1,NP-252340.1,ZP-00022353.1,NP-355549.1,NP-420724.1.,ZP-00085169.1,EAA17616.1,NP-273242.1,NP-219574.1NP-387094.1,NP-296721.1,ZP-00004209.1,NP-823739.1,NP-282934.1,BAA77848.1,NP-660577.1,NP-760741.1,NP-641750.1,NP-636741.1,NP-829309.1,NP-298338.1,NP-444964.1,NP-717246.1,NP-224545.1,ZP-00038451.1,DXR_KITGR,NP-778563.1。異戊烯基-二磷酸[δ]-異構酶基因的例子是:編碼來自巴勒斯坦側金盞花的異戊烯基-二磷酸[δ]-異構酶的核酸,以及進一步來自其他生物的具有如下登錄號的異戊烯基-二磷酸[δ]-異構酶基因:Q38929,O48964,Q39472,Q13907,O35586,P58044,O42641,O35760,Q10132,P15496,Q9YB30,Q8YNH4,Q42553,O27997,P50740,O51627,O48965,Q8KFR5,Q39471,Q39664, Q9RVE2,Q01335,Q9HHE4,Q9BXS1,Q9KWF6,Q9CIF5,Q88WB6,Q92BX2,Q8Y7A5,Q8TT35Q9KK75,Q8NN99,Q8XD58,Q7FE75,Q46822,Q9HP40,P72002,P26173,Q9Z5D3,Q8Z3X9,Q8ZM82,Q9X7Q6,O13504,Q9HFW8,Q8NJL9,Q9UUQ1,Q9NH02,Q9M6K9,Q9M6K5,Q9FXR6,O081691,Q9S7C4,Q8S3L8,Q9M592,Q9M6K3,Q9M6K7,Q9FV48,Q9LLB6,Q9AVJ1,Q9AVG8,Q9M6K6,Q9AVJ5,Q9M6K2,Q9AYS5,Q9M6K8,Q9AVG7,Q8S3L7,Q8W250,Q94IE1,Q9AVI8,Q9AYS6,Q9SAY0,Q9M6K4,Q8GVZ0,Q84RZ8,Q8KZ12,Q8KZ66,Q8FND7,Q88QC9,Q8BFZ6,BAC26382,CAD94476。香葉基-二磷酸合酶基因的例子是:編碼來自擬南芥的香葉基-二磷酸合酶的核酸,以及進一步來自其他生物的具有如下登錄號的香葉基-二磷酸合酶基因:Q9FT89,Q8LKJ2,Q9FSW8,Q8LKJ3,Q9SBR3,Q9SBR4,Q9FET8,Q7LKJ1,Q84LG1,Q9JK86。法呢基-二磷酸合酶基因的例子是:編碼來自擬南芥的法呢基-二磷酸合酶的核酸,以及進一步來自其他生物的具有如下登錄號的法呢基-二磷酸合酶基因:P53799,P37268,Q02769,Q09152,P49351,O24241,Q43315,P49352,O24242,P49350,P08836,P14324,P49349,P08524,O66952,Q082911P54383,Q45220,P57537,Q8K9A0,P22939,P45204,O66126,P55539,Q9SWH9,Q9AVI7,Q9FRX2,Q9AYS7,Q94IE8,Q9FXR9,Q9ZWF6,Q9FXR8,Q9AR37,O50009,Q94IE9,Q8RVK7,Q8RVQ7,O04882,Q93RA8,Q93RB0,Q93KB4,Q93RB5,Q93RB3,O93RB1,Q93RB2,Q920E5。香葉基-香葉基-二磷酸合酶基因的例子是:編碼來自白芥(Sinapisalba)的香葉基-香葉基-二磷酸合酶的核酸,以及進一步來自其他生物的香葉基-香葉基-二磷酸合酶基因。八氫番茄紅素合酶基因的例子是:編碼來自噬夏孢歐文氏菌的八氫番茄紅素合酶的核酸,以及進一步來自其他生物的八氫番茄紅素合酶基因。八氫番茄紅素去飽和酶基因的例子是:編碼來自噬夏孢歐文氏菌的八氫番茄紅素去飽和酶的核酸,以及進一步來自其他生物的八氫番茄紅素去飽和酶基因。ζ-胡蘿蔔素去飽和酶基因的例子是:編碼來自洋水仙的ζ-胡蘿蔔素去飽和酶的核酸,以及進一步來自其他生物的ζ-胡蘿蔔素去飽和酶基因。crtISO基因的例子是:編碼來自番茄的crtISO的核酸,以及進一步來自其他生物的crtISO基因。本發明還涉及經遺傳修飾的耶氏酵母屬微生物,其中所述遺傳修飾導致與野生型相比提高的表達速率或引起至少一個基因的表達,並且所述表達是由本發明所述啟動子引起的。在本發明的經遺傳修飾的耶氏酵母屬生物的一個優選實施方案中,本發明的啟動子在所述生物體中對所述基因的表達調控是通過如下實現的:a)將一個或多個上述重組核酸分子引入所述微生物的基因組中,使得所述微生物一個或多個內源基因的表達在引入的重組核酸分子控制下發生;b)將一個或多個基因引入所述微生物的基因組中,使得一個或多個引入的基因的表達在上述重組核酸分子控制下發生,所述重組核酸分子對於宿主微生物是內源的;或c)將一個或多個核酸構建體引入所述微生物中,其中所述一個或多個核酸構建體包含至少一個上述重組核酸分子,且此一個或多個重組核酸分子與一個或多個要表達的基因是功能上相連的。在一個實施方案中,可以在染色體內或染色體外將所述核酸構建體整合入根據特徵c)的耶氏酵母屬的微生物中。本發明的優選的啟動子和優選的要表達的基因(效果基因)如上所述。具有效果基因的提高的或引發的表達速率的經遺傳修飾的耶氏酵母屬的微生物的生產在下文通過例示方式進行描述。在遺傳修飾組合的情況下,可以進行單一轉化或通過多個構建體轉化。優選通過用核酸構建體轉化初始生物來產生轉基因生物,所述核酸構建體包含上述本發明的啟動子的至少一個,其與要表達的效果基因是功能上相連的,若適當的話,其還與進一步的調控信號是功能上相連的。核酸構建體(其中本發明的啟動子和效果基因是功能上相連的)在下文也稱為表達盒。表達盒可以包含進一步的調控信號,其是控制效果基因在宿主細胞中表達的調控核酸序列。在一個優選實施方案中,表達盒在編碼序列的上遊(即其5』端)包含本發明的至少一個啟動子,和多聚腺苷酸化信號,並且如果適當的話,還在下遊(即在3』端)包含進一步的調控元件,所述調控元件可操作地連接至位於其間的(lyinginbetween)對於上述基因的至少一個的效果基因的 編碼序列。可操作連接意指啟動子,編碼基因,終止子,適當的話以及進一步的調控元件的連續排列,該排列的方式使得每一個調控元件能在編碼基因的表達過程中完成其意欲的功能。對於生物優選的核酸構建體、表達盒和載體及用於生產轉基因生物的方法,以及轉基因的耶氏酵母屬生物本身,描述於WO2006/102342中,並通過如下實施例加以描述。本發明的核酸可以通過合成產生或天然地獲得,或是包含了合成和天然核酸成分的混合物,並由來自不同生物的多種異源基因區段組成。針對生物偏好的密碼子,如上所述,對合成的核苷酸序列給予了偏好性。這些生物所偏好的密碼子可以從大多數感興趣的生物種中表達的最高蛋白質頻率的密碼子來確定。在表達盒的製備中,可能操縱多種DNA片段,從而獲得容易以正確方向讀取且配有正確讀框的核苷酸序列。可以將銜接頭或接頭附接至片段,以共同加入DNA片段中。用包含一個或多個用於插入該序列的限制位點的接頭或多聚接頭,是很方便地能以轉錄的方向來提供啟動子和終止子區的。通常,接頭具有1-10個限制位點,大多數情況下1-8個、優選2-6個限制位點。一般而言,調控區內的接頭具有小於100bp,經常小於60bp,但至少為5bp的大小。啟動子對於宿主生物而言可以是天然的(或同源的)也可以是外來的(或異源的)。表達盒優選以5'-3'轉錄方向包含啟動子、編碼核酸序列或核酸構建體以及轉錄終止區。不同的終止區可以根據需要互換。特別優選的效果基因是選自下組的那些:編碼酮酶的核酸、編碼[β]-羥化酶的核酸、編碼[β]-環化酶的核酸、編碼[ε]-環化酶的核酸、編碼環氧酶的核酸、編碼HMG-CoA還原酶的核酸、編碼(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶的核酸、編碼1-脫氧-D-木糖-5-磷酸合酶的核酸、編碼1-脫氧-D-木糖-5-磷酸還原異構酶的核酸、編碼異戊烯基-二磷酸[δ]-異構酶的核酸、編碼香葉基-二磷酸合酶的核酸、編碼法呢基-二磷酸合酶的核酸、編碼香葉基-香葉基-二磷酸合酶的核酸、編碼八氫番茄紅素合酶的核酸、編碼八氫番茄紅素去飽和酶的核酸、編碼前八氫番茄紅素合酶的核酸、和編碼ζ-胡蘿蔔素去飽和酶的核酸。藉助上述本發明的方法可能的是,在耶氏酵母屬的上述本發明的經遺傳修飾的生物中通過本發明的啟動子將代謝途徑調控至特定的生物合成產物。出於這個目的,例如,通過引發或提高此生物合成途徑的基因的轉錄速率或表達速率以增加的酶量來增強該種導致特定生物合成產物的代謝途徑,其中所述增加的酶量導致合意的生物合成途徑的這些酶的總活性增加,並因而導致指向合意的生物合成產物的代謝流增強。取決於合意的生物合成產物,需要提高或降低多個基因的轉錄速率或表達速率。改變多個基因的轉錄速率或表達速率通常是有利的,即,提高基因的組合的轉錄速率或表達速率和/或降低基因的組合的轉錄速率或表達速率。在本發明的經遺傳修飾的生物中,至少一個提高的或引發的基因表達速率可以歸因於本發明的啟動子。此外,在經遺傳修飾的生物中其他基因的額外改變的(即額外提高或額外降低的)表達速率可以(但不需要)來源於本發明的啟動子。因此,本發明涉及用於通過培養耶氏酵母屬的本發明的經遺傳修飾的生物來生產生物合成產物的方法。具體而言,本發明涉及用於通過培養耶氏酵母屬的本發明的經遺傳修飾的生物來生產類胡蘿蔔素的方法,其中要表達的基因選自下組:編碼酮酶的核酸、編碼[β]-羥化酶的核酸、編碼[β]-環化酶的核酸、編碼[ε]-環化酶的核酸、編碼環氧酶的核酸、編碼HMG-CoA還原酶的核酸、編碼(E)-4-羥基-3-甲基丁-2-烯基-二磷酸還原酶的核酸、編碼1-脫氧-D-木糖-5-磷酸合酶的核酸、編碼1-脫氧-D-木糖-5-磷酸還原異構酶的核酸、編碼異戊烯基-二磷酸[δ]-異構酶的核酸、編碼香葉基-二磷酸合酶的核酸、編碼法呢基-二磷酸合酶的核酸、編碼香葉基-香葉基-二磷酸合酶的核酸、編碼八氫番茄紅素合酶的核酸、編碼八氫番茄紅素去飽和酶的核酸、編碼前八氫番茄紅素合酶的核酸、和編碼ζ-胡蘿蔔素去飽和酶的核酸。類胡蘿蔔素優選為選自下組:八氫番茄紅素、六氫番茄紅素、番茄紅素、葉黃素、β-胡蘿蔔素、玉米黃素、蝦青素、角黃素、海膽酮、3-羥基海膽酮、3』-羥基海膽酮、金盞花紅素、堇黃素和金盞花黃素。此外,經遺傳修飾的生物還可用於生產含有生物合成產物(尤其是類胡蘿蔔素,尤其是酮式類胡蘿蔔素,尤其是蝦青素)的提取物,和/或用於生產動物和人食物補充劑,以及化妝品和藥物。耶氏酵母屬的經遺傳修飾的生物與野生型相比具有增加的合意的生物合成產物,尤其是類胡蘿蔔素,尤其是酮式類胡蘿蔔素,尤其是蝦青素的含量。在優選的實施方案中,本發明的酵母啟動子來源於解脂耶氏酵母菌株。目前預期,本發明的酵母啟動子(即同功的酵母啟動子)可以獲得自其他微生物。例如,酵母啟動子可以來源於其他酵母菌株,如釀酒酵母(Saccharomycescerevisiae)的菌株。在另一個方面,本發明提供包含本發明的酵母啟動子的重組表達載體。本發明的表達載體可為便利地用於重組DNA步驟的任何表達載體,並且載體的選擇通常取決於要將其引入的宿主細胞。用於分別連接編碼目的蛋白的DNA序列、酵母啟動子和終止子的步驟,以及將它們插入合適的載體的步驟是本領域技術人員已知的。在又一個方面,本發明提供包含本發明的重組表達載體的宿主細胞。優選地,本發明的宿主細胞是真核細胞,尤其是酵母細胞。此類酵母宿主細胞的例子包括但不限於酵母屬(Saccharomyces)菌株,尤其是釀酒酵母、克魯弗酵母(Saccharomyceskluyveri)、或葡萄汁酵母(Saccharomycesuvarum)、裂殖酵母屬菌種(Schizosaccharomycessp.),如粟酒裂殖酵母(Schizosaccharomycespombe)的菌株、漢遜酵母屬菌種(Hansenulasp.)的菌株、畢赤酵母屬菌種(Pichiasp.)、耶氏酵母屬菌種,如解脂耶氏酵母、或克魯維酵母屬菌種(Kluyveromycessp.),如乳酸克魯維酵母(Kluyveromyceslactis)。在優選的實施方案中,解脂耶氏酵母的菌株是適於本發明的宿主。實施例如下實施例對本發明進行闡釋。下表1描述了用於下文範例中的一些解脂耶氏酵母菌株:表1:解脂耶氏酵母菌株下表2描述了用於下文範例中的一些質粒表2:質粒質粒主鏈/骨架插入物pMB6056pMB6052(HygRtef1P-xprT)crtZpMB6504pMB6056hspPpMB6509pMB6056hypPpMB6779pMB6149(LEU2lacZ)hspPpMB6781pMB6149hypP通過引入異源基因在內源性TEF1啟動子的控制下(對照),構建了耶氏酵母株ML2461和ML6804。GGS基因和截短的HMG基因(「HMG-tr」)分別來源於與天然香葉基-香葉基焦磷酸合酶和羥基甲基戊二醯-CoA還原酶基因對應的耶氏酵母屬序列。carRP和carB基因來源於卷枝毛黴(Mucorcircinelloides),並且它們分別編碼雙功能性的八氫番茄紅素合酶/番茄紅素環化酶和八氫番茄紅素脫氫酶。合成了crtW基因以編碼百慕達細小棒菌(Parvularculabermudensis)的胡蘿蔔素酮酶。合成了crtZ基因以編碼粉末腸桿菌(Enterobacterpulveris)的胡蘿蔔素羥化酶。這些基因有時(但不總是)與營養缺陷型標誌物(URA3,LEU2,URA2,LYS1,ADE1)或loxP位點、HygR或NatR標誌物的殘餘聯繫起來。此處所述的所有基礎分子生物學及DNA操作步驟通常是根據Sambrook等或Ausubel等(SambrookJ,FritschEF,ManiatisT(編).1989.MolecularCloning:ALaboratoryManual.ColdSpringHarborLaboratoryPress:NewYork;AusubelFM,BrentR,KingstonRE,MooreDD,SeidmanJG,SmithJA,StruhlK(編).1998.CurrentProtocolsinMolecularBiology.Wiley:NewYork)進行的。實施例1:生產pM6504(HygRHSPp-crtZ),其編碼驅動胡蘿蔔素羥化酶基因crtZ的HSP啟動子用MO8250(5』-CACACAAAGCTTGGTACCAGATAGTGCAATCACATGTTGCTAC)和MO8251(5』-CACACATTTTGTGGCTAGCATGTAGTATAAGTGTGTGTGTTGG)從製備自解脂耶氏酵母ATCC201249的gDNA擴增了對應於YALI0D20526g上遊的基因內區域的SEQIDNO:1中指定的序列。用NheI和HindIII來切割此1kb片段,並連接至用NheI和HindIII切割的pMB6056(TEF1p-crtZHygR)以產生pMB6504(HSPp-crtZHygR)。實施例2:生產pM6509(HygRHSPp-crtZ),其編碼驅動胡蘿蔔素羥化酶基因crtZ的來自解脂耶氏酵母的HYP啟動子用MO8254(5』-CACACAAAGCTTGGTACCAGATCGCGGTCAGAAGGGGCAG)和MO8255(5』-CACACATTTTGTGGCTAGCATGTGAGAAAGTAGTTTTGAGATGG)從製備自解脂耶氏酵母ATCC201249的gDNA擴增了對應於YALI0D09889g上遊的基因內區域的SEQIDNO:2中指定的序列。用NheI和HindIII來切割此1kb片段,並連接至用NheI和HindIII切割的pMB6056(TEF1p-crtZHygR)以產生pMB6509(HYPp-crtZHygR)。實施例3:生產玉米黃素用NotI切割pMB6056,pMB6504和pMB6509並獨立地轉染至ML2461(一種β胡蘿蔔素生產者)中,以及ML6804(一種角黃素生產者)中。將轉化體在補充100mg/L潮黴素B的YPD上進行選擇。將它們在30℃溫育48小時。挑取菌落至補充100mg/L潮黴素B的YPD瓊脂。使這些轉化體的培養物在2mL圓底24孔板中的800μLYPD中於30℃生長72小時,並在INFORSMultitron中以800rpm振蕩。為了提取,取樣250μL至2mLEppendorfSafe-LockTM管(022363352)中。向每個樣品添加約600μL的0.5mm直徑氧化鋯/二氧化矽珠(BioSpecProducts目錄號11079105z)。在ReschMM300珠磨器(設置為20)中於庚烷:乙酸乙酯(1:1v:v)中於4℃將細胞破碎5分鐘,並離心5分鐘。進行多輪提取直至提取物無色,指示樣品中類胡蘿蔔素已耗盡。在破碎之後將提取物匯集,蒸發、重懸於己烷:乙酸乙酯(1:1v:v)中並通過HPLC分析。圖1顯示了在搖瓶中用驅動crtZ表達的HSP或HYP來轉化的ML2461中的類胡蘿蔔素水平,描繪為HPLC峰面積。由於耶氏酵母屬中的插入是隨機的,因此染色體定位隨轉化體而異。為了將位置效應降至最小,對HSP和HYP分別測定了10個和5個轉化體並取平均值。實施例4:生產蝦青素培養來源於ML6804轉化體的培養物,取樣並如上文所述提取。圖2顯示了用驅動crtZ表達的HSP或HYP啟動子來轉化的ML6804中的類胡蘿蔔素水平, 描繪為HPLC峰面積。顯示了搖瓶中對於HSP和HYP各12個轉化體的平均表達。實施例5:蝦青素生產者的發酵罐中的基因表達為了檢查上述異源構建體在發酵罐條件下的表達,將實施例4中產生的產蝦青素的菌株用分批補料工藝培養於發酵罐中,所述培養在3L實驗室規模(bench-scale)發酵罐中進行。初始批次階段(initialbatchphase)培養基含有作為主要碳源的10%大豆油(vol:vol)。在批式生長期過程中,生物質水平(幹細胞重量)達到最大,並且耶氏酵母細胞積累大的內部脂質體。在消耗了初始批次後,觀察到溶解氧水平在發酵罐中的快速上升。在此時,開始進料大豆油,控制補料速度以將發酵罐溶解氧水平維持在20%的飽和度。在圖3中指示的時間點處取樣25μL的等分試樣並如上文所述提取類胡蘿蔔素。對於RNA分析,對發酵罐一次取一個樣以將從取樣到冷凍的時間最小化。取出樣品,並立即置於冰上。一式三份取等分試樣,並在2mLEppendorfSafe-LockTM管中於13000rpm離心45秒。棄上清,並將所述樣品置於乾冰上,直至所有發酵罐均已取樣。將樣品儲藏在-80℃。用QiagenRNeasyMiniRNA提取試劑盒(目錄號74106)採用YeastIII規程伴隨珠打(beadbeating)來提取RNA。向每個樣品添加約600μL的0.5mm直徑氧化鋯/二氧化矽珠(BioSpecProducts目錄號11079105z)。向樣品添加600μLBufferRLT及β-ME(10μLβ-ME/1mLRLT)的等分試樣。在ReschMM300珠磨器(設置為20)中於4℃將細胞破碎5分鐘。以最高速離心樣品10秒並將上清轉移至新管。將樣品再次以最高速離心2分鐘並將裂解液轉移至新管。向裂解液添加350μL的70%乙醇的等分試樣,並通過上下移取進行混合。將700μL的樣品的等分試樣轉移至RNeasymini柱並於13000rpm離心15秒。將流過物(flow-through)棄去。將350μLBufferRW1的等分試樣添加至該柱,在13000rpm將其離心15秒。將10μLDnaseI儲液的等分試樣添加至70μL的BufferRDD並通過顛倒管體進行混合。將80μLDNaseI溶液添加至RNeasy二氧化矽-凝膠膜並在室溫溫育15分鐘。將350μLBufferRW1的等分試樣添加至該柱並在13000rpm將其離心15秒。將流過物棄去。將RNeasy柱轉移至新的2mL收集管。為了洗滌柱子,添加500μLBufferRPE(含乙醇)的等分試樣,並在13000rpm將其離心15秒。將流過物棄去,並重複之前的步驟。將RNeasy柱置於新的2mL收集管中並於 13000rpm離心1分鐘以將膜乾燥。將RNeasy柱轉移至新的1.5mL收集管。將50μL的無RNA酶水的等分試樣添加至膜,並在室溫溫育1分鐘。以13000rpm將柱離心以洗脫。如上所述在Agilent2100生物分析儀上用AgilentRNAPico(Agilent#5067-1513)試劑盒對RNA進行定量並測試其質量。用VILO試劑盒(Invitrogen目錄號11754-250)根據製造商的指導來合成第一鏈cDNA。簡言之,將250ng的RNA在20μL反應中與4μL5xVILO反應混合劑(ReactionMix)、2μL10xSuperScript酶混合劑及水一起溫育。將反應在25℃溫育10分鐘,42℃溫育2小時並在85℃溫育5分鐘。將cDNA稀釋至0.7ng/μL然後進行qPCR。用PrimerQuest軟體(可免費獲取自IDT網站)設計,並通過IDT合成用於qPCR的引物。表3:用於qPCR的引物在StratageneMx3000P熱循環儀中用BrilliantIIIUltra-FastSYBERQPCR主混合物(MasterMix)(Agilent目錄號600883)來進行qPCR。運行了幾個20μL的反應。主混合物設置為含有除cDNA之外的全部成分,並向每個反應添加15μL等分試樣以及5μL的cDNA。向每個反應添加10μL2xSYBERMix,0.3μL的500x稀釋的參考染料(ReferenceDye),0.7μL水和4μL的引物混合物(正向和反向引物各2μM)。還用ACT1引物組和0.7ng/μLRNA作為模板運行了無逆轉錄酶反應(NRT)。對每個平板運行了無模板(NTC)反應,其用5μL的水取代了cDNA。如果一個實驗運行多個平板,則還包括平板間校準物(interplatecalibrator,IPC))。對於IPC,選擇單個模板並用ACT1引物組 在所有平板中一式三份運行。所有反應均一式三份運行。選擇ACT1以對mRNA量進行標準化,並將Tef1α(一個組成型看家基因)用於倍數表達計算。計算dCt以去除變異(Avg樣品-AvgACT1)。計算ddCt以標準化至標準物,在本研究中為TEF1α,其為一種強表達的內源性基因。(2^(dCt樣品-dCtTEF))。圖3顯示了由產生的蝦青素水平反映的crtZ表達的水平。圖4顯示了在3個時間點處來自與圖3相同發酵罐的相對crtZRNA表達。實施例6:生產β-半乳糖苷酶為了評估來自HYP和HSP啟動子的基因表達,將每個啟動子功能性地融合至lacZ報告基因。用NheI和HindIII切割質粒pMB6149,並將實施例1和2中所述合適的1kbNheI-HindIII啟動子片段插入以產生pMB6779(HSPp-lacZ)和pMB6781(HYPp-lacZ)。將來源於ML326的Leu+轉化體的培養物培養於補充了尿嘧啶和賴氨酸的基本培養基中過夜,然後1/50稀釋於10mLYPD中,並生長至2x107細胞/mL。將細胞沉澱並重懸於0.5mL破裂緩衝液(BreakingBuffer)(0.1MTrispH8,20%甘油v/v,1mMDTT)中。將0.25mL的等分試樣轉移至新鮮的管中,並添加12.5uLPMSF溶液(40mM溶於95%乙醇中)。將氧化鋯/二氧化矽珠(BioSpecProducts目錄號11079105z)添加至彎液面,並將管渦旋10分鐘。將上清轉移至新鮮的管。在4℃將溶液以最大速度離心15分鐘。將950μL的Z緩衝液(16g/LNa2HPO4-7H2O,5.5g/LNaH2PO4-H2O,0.75g/LKCl,0.25g/LMgSO4·7H2O,2.7mL/Lβ-巰基乙醇)和50μL提取物的等分試樣組合於玻璃管中並置於28℃水浴中。通過添加0.2mLONPG溶液(4mg/mLO-硝基苯基B-D-半乳糖苷於Z緩衝液中)來開始反應。允許反應繼續直至顯現中黃色(mediumyellowcolor),並通過添加0.5mL1MNa4CO3來終止反應。記錄每個反應的時間並確定OD420。通過BioRad蛋白測定法來確定總細胞蛋白。β-半乳糖苷酶單位=((OD420)(378)/((以分鐘計的時間)(以mL計的提取物體積)(蛋白mg/mL))圖5顯示了搖瓶中分別用HSP或HYP(其驅動單拷貝lacZ的表達)的4株和5株中β-半乳糖苷酶生產。當前第1頁1 2 3