改良的重編程方法和細胞培養平臺與流程
2023-05-26 02:13:17 3
本專利申請根據35 U.S.C.§119(e)要求於2014年3月4日提交的美國臨時申請號61/947,979的利益,所述美國臨時申請以引用的方式全文併入本文。
關於序列表的陳述
與本專利申請相關的序列表以文本形式代替文件拷貝提供,並且在此以引用的方式併入說明書內。含有序列表的文本文件的名稱為FATE_122_01WO_ST25.txt。文本文件是8KB,於2015年3月4日創建,並且與說明書提交同時經由EFS-Web電子提交。
技術領域
本發明一般涉及用於製造多能細胞的組合物和方法。特別地,本發明涉及用於製造具有基態多能性的多能細胞的改良的培養平臺。
背景技術:
當今的基於多能幹細胞的疾病和毒理學篩選努力以及明日的自體/同種異體多能幹細胞療法將需要人胚胎幹細胞(hESC)和人誘導多能幹細胞(hiPSC)的細胞系生成和擴增的穩健、可重現方法。hiPSC已通過經由基因組整合逆轉錄病毒和慢病毒表達系統引入的多能性因子的異位表達生成。消除儘可能多的整合事件的努力已包括用小分子抑制劑代替許多重編程因子。另外,非整合方法已證明是無效和費力的,從而需要另外的重編程因子或在使所有體細胞重編程中是無效的(Lee等人,2013)。
與多能幹細胞培養相關的幾個挑戰仍有待解決,以允許細胞變得適合於未來的工業和臨床應用。在最常用的常規培養系統中,hESC和hiPSC在作為塊傳代時維持在飼養細胞上,以防止廣泛細胞死亡和基因組異常(Thomson等人,1998)。在無飼養者(FF)環境中不能單細胞培養hiPSC嚴重限制潛在的工業規模篩選或細胞療法應用(Skottman等人,2007;Valamehr等人,2011)。另外,關於改良hiPSC的近期努力已集中於慢病毒衍生的hiPSC,所述hiPSC並非是無轉基因的,從而限制此類努力的治療關聯性。
仍有待成功解決的另一個挑戰,即基因組修飾的缺乏,是培養中的人多能幹細胞的自發分化的傾向(Pera和Trounson,2004;Sathananthan和Trounson,2005;Valamehr等人,2011)。
hESC和hiPSC中的研究已得到描述,但需要多能性基因的連續異位表達以維持基態,從而導致基因組修飾的人多能幹細胞(Hanna等人,2010a),所述人多能幹細胞不適合於工業和臨床級別的多能細胞。
相應地,關於人多能細胞產物的高流通量、無轉基因或足跡生成的組合物和方法的不存在迄今被證明是在未來的多能幹細胞療法的發展和商業化中的重大障礙。
技術實現要素:
本發明一般提供了改良的細胞培養平臺。
在各個實施例中,本發明部分考慮了包含下述的組合物:(a)Wnt途徑激動劑;(b)MEK抑制劑;和(c)ROCK抑制劑,其中所述組合物不包含TGFβR抑制劑。
在特定實施例中,Wnt途徑激動劑是GSK3抑制劑。
在某些實施例中,GSK3抑制劑是CHIR99021或BIO。
在另外的實施例中,MEK抑制劑是PD98059或PD032901。
在進一步的實施例中,ROCK抑制劑是thiazovivin或Y27632。
在一些實施例中,GSK3抑制劑是CHIR99021;MEK抑制劑是PD032901;並且ROCK抑制劑是thiazovivin。
在某些實施例中,前述組合物中的任一種還包含bFGF或LIF。
在進一步的實施例中,前述組合物中的任一種還包含bFGF和LIF。
在各個實施例中,本發明提供了包含前述組合物中的任一種的培養基,其中所述培養基不包含TGFβR抑制劑。
在一些實施例中,培養一個或多個多能細胞的方法包括在根據前述細胞培養基中的任一種的細胞培養基中培養所述一個或多個多能細胞。
在另外的實施例中,一個或多個多能細胞是胚胎幹細胞(ESC)或誘導多能幹細胞(iPSC)。
在特定實施例中,一個或多個多能細胞是iPSC。
在某些實施例中,組合物包含多能細胞群。
在進一步的實施例中,多能細胞群是多能細胞的同質群。
在特定實施例中,多能細胞群的至少95%表達SSEA4-FITC和TRA1-81或TRA1-60。
在一些實施例中,多能細胞群的至多5%表達α-平滑肌肌動蛋白(SMA)、TUJ1或FoxA2。
在特定實施例中,多能細胞先前在包含TGFβR抑制劑的細胞培養基中進行培養。
在另外的實施例中,在細胞培養基中培養多能細胞減少培養細胞的自發分化。
在一個實施例中,與一種或多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的多能細胞中的表達相比較,一種或多種分化標記物基因在培養細胞中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在另一個實施例中,一種或多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在另外一個實施例中,與兩種或更多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的多能細胞中的表達相比較,所述兩種或更多種分化標記物基因在培養細胞中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在另外一個實施例中,兩種或更多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在一個特定實施例中,與三種或更多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的多能細胞中的表達相比較,所述三種或更多種分化標記物基因在培養細胞中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在某些實施例中,三種或更多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在另外的實施例中,與五種或更多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的多能細胞中的表達相比較,所述五種或更多種分化標記物基因在培養細胞中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在一個進一步的實施例中,五種或更多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在某些實施例中,在細胞培養基中培養多能細胞維持或誘導多能性的基態。
在特定實施例中,一個或多個多能細胞的多能性的基態維持至少5代。
在某些特定實施例中,一個或多個多能細胞的多能性的基態維持至少10代。
在進一步的特定實施例中,一個或多個多能細胞的多能性的基態維持至少50代。
在另外的特定實施例中,一個或多個多能細胞的多能性的基態維持至少100代。
在各個實施例中,前述方法還包括在傳代期間解離一個或多個多能細胞。
在某些實施例中,一個或多個多能細胞的活力在傳代期間得到維持。
在某些特定實施例中,一個或多個多能細胞包含正常核型。
在某些另外的實施例中,一個或多個多能細胞在無飼養者環境中進行培養。
在某些進一步的實施例中,一個或多個多能細胞的基因組穩定性維持至少10代。
在某些相關實施例中,一個或多個多能細胞的基因組穩定性維持至少50代。
在某些其他實施例中,一個或多個多能細胞的基因組穩定性維持至少100代。
在各個實施例中,本發明部分考慮了使多能細胞適應無飼養者培養的方法,所述方法包括:(a)分離在飼養細胞的存在下培養的一個或多個多能細胞;(b)在化學成分確定的細胞培養基中培養一個或多個多能細胞,所述細胞培養基包含:Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和ROCK抑制劑,其中所述培養基不包含TGFβR抑制劑。
在各個特定實施例中,本發明部分考慮了培養作為單細胞酶促傳代的多能細胞的方法,所述方法包括:(a)酶促處理一個或多個多能細胞,以傳代單個多能細胞;(b)在無飼養者環境中培養單個多能細胞;(c)在化學成分確定的細胞培養基中培養單個多能細胞,所述細胞培養基包含:Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和ROCK抑制劑,其中所述培養基不包含TGFβR抑制劑。
在各個某些實施例中,本發明部分考慮了減少一個或多個多能細胞的自發分化的方法,所述方法包括:(a)在無飼養者環境中培養一個或多個多能細胞;(b)在化學成分確定的細胞培養基中培養一個或多個多能細胞,所述細胞培養基包含:Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和ROCK抑制劑,其中所述培養基不包含TGFβR抑制劑。
在各個另外的實施例中,本發明部分考慮了製造誘導多能幹細胞(iPSC)的方法,所述方法包括:(a)獲得一個或多個非多能細胞;(b)使一個或多個非多能細胞重編程為多能狀態;(c)在不包含TGFβR抑制劑的細胞培養基中培養多能細胞,由此產生iPSC。
在特定實施例中,一個或多個非多能細胞包含體細胞。
在一些實施例中,一個或多個非多能細胞包含成人幹細胞。
在某些實施例中,一個或多個非多能細胞通過增加細胞中的內源OCT4表達而重編程為多能狀態。
在進一步的實施例中,使一個或多個非多能細胞重編程為多能狀態包括將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、ECAT1、UTF1、ESRRB和SV40LT。
在另外的實施例中,使一個或多個非多能細胞重編程為多能狀態包括將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、ECAT1、UTF1、ESRRB和SV40LT。
在某些實施例中,使一個或多個非多能細胞重編程為多能狀態包括將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、SOX2和NANOG。
在某些實施例中,使一個或多個非多能細胞重編程為多能狀態包括將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、NANOG、ECAT1、UTF1和ESRRB。
在某些實施例中,使一個或多個非多能細胞重編程為多能狀態包括將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、ECAT1和UTF1。
在特定實施例中,一種或多種多核苷酸是慢病毒載體。
在一些實施例中,一種或多種多核苷酸是附加型載體。
在相關特定實施例中,細胞培養基包含Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和ROCK抑制劑。
在進一步的特定實施例中,使一個或多個非多能細胞重編程包括使一個或多個非多能細胞與下述接觸:Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和TGFβR抑制劑,以及任選的ROCK抑制劑。
在另外的實施例中,iPSC包含iPSC群。
在特定實施例中,iPSC群是iPSC的同質群。
在特定實施例中,iPSC群的至少95%表達SSEA4和TRA1-81或TRA1-60。
在某些實施例中,多能細胞群的至多5%表達α-平滑肌肌動蛋白(SMA)、TUJ1或FoxA2。
在特定實施例中,在細胞培養基中培養多能細胞減少自發分化,或者維持或誘導多能性的基態。
在另外的實施例中,與一種或多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的iPSC中的表達相比較,一種或多種、兩種或更多種、三種或更多種、四種或更多種、或者五種或更多種分化標記物基因在所述iPSC中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在特定實施例中,一種或多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在某些實施例中,自發分化中的減少維持至少5代。
在其他實施例中,自發分化中的減少維持至少10代。
在特定實施例中,自發分化中的減少維持至少50代。
在另外的實施例中,自發分化中的減少維持至少100代。
在進一步的實施例中,前述方法包括在傳代期間解離iPSC。
在另外的實施例中,iPSC的活力在傳代期間得到維持。
在某些實施例中,iPSC包含正常核型。
在其他實施例中,iPSC在無飼養者環境中進行培養。
在特定實施例中,iPSC的基因組穩定性維持至少10代。
在另外的實施例中,iPSC的基因組穩定性維持至少50代。
在特定的另外實施例中,iPSC的基因組穩定性維持至少100代。
在各個實施例中,本發明部分提供了根據前述實施例中任何一個產生的包含基態多能性的誘導多能幹細胞(iPSC)。
在各個某些實施例中,本發明部分提供了包含基態多能性的誘導多能幹細胞(iPSC),其中所述iPSC不包含編碼重編程因子多肽的外源引入的多核苷酸。
在各個特定實施例中,本發明部分提供了製造誘導多能幹細胞(iPSC)的方法,所述方法包括:a)獲得一個或多個多能幹細胞;(b)在不包含TGFβR抑制劑的細胞培養基中培養一個或多個多能幹細胞,由此產生基態iPSC。
在某些實施例中,一個或多個iPSC包含重編程的體細胞。
在另外的實施例中,一個或多個iPSC包含重編程的成人幹細胞。
在其他實施例中,一個或多個iPSC通過增加一個或多個iPSC中的內源OCT4表達而重編程為多能狀態。
在特定實施例中,一個或多個iPSC通過將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸引入一個或多個非多能細胞內進行重編程:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、ECAT1、UTF1、ESRRB和SV40LT。
在某些特定實施例中,一個或多個iPSC通過引入編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸進行重編程:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、ECAT1、UTF1、ESRRB和SV40LT。
在另外的實施例中,一個或多個iPSC通過將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入所述一個或多個非多能細胞內進行重編程:OCT4、SOX2和NANOG。
在另外的實施例中,一個或多個iPSC通過將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入所述一個或多個非多能細胞內進行重編程:OCT4、NANOG、ECAT1、UTF1和ESRRB。
在另一個實施例中,一個或多個iPSC通過將編碼選自下述的重編程因子的一個或多個拷貝的一種或多種多核苷酸引入所述一個或多個非多能細胞內進行重編程:OCT4、ECAT1和UTF1。
在各個實施例中,慢病毒載體包含一種或多種多核苷酸。
在特定實施例中,附加型載體包含一種或多種多核苷酸。
在其他實施例中,細胞培養基包含Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和ROCK抑制劑。
在某些實施例中,獲得一個或多個iPSC包括使一個或多個非多能細胞或部分多能細胞與下述接觸:Wnt途徑激動劑,任選地其中所述Wnt途徑激動劑是GSK3抑制劑;MEK抑制劑;和TGFβR抑制劑,以及任選的ROCK抑制劑,以產生一個或多個iPSC。
在另外的實施例中,iPSC包含iPSC群。
在另外的實施例中,iPSC群是iPSC的同質群。
在特定實施例中,iPSC群的至少95%表達SSEA4和TRA1-81或TRA1-60。
在特定實施例中,一個或多個iPSC通過使多能細胞群重編程而獲得,其中所述多能細胞群的至多5%表達α-平滑肌肌動蛋白(SMA)、TUJ1或FoxA2。
在進一步的實施例中,前述方法包括在細胞培養基中培養一個或多個iPSC減少自發分化,或者維持或誘導多能性的基態。
在某些實施例中,具有減少的自發分化的iPSC包含基因表達,其中與一種或多種分化標記物基因在包含TGFβR抑制劑的培養基中培養的iPSC中的表達相比較,一種或多種、兩種或更多種、三種或更多種、四種或更多種、或者五種或更多種分化標記物基因在iPSC中的表達減少至少約10%、20%、25%、30%、40%、50%、60%、70%、80%或90%,其中所述分化標記物基因選自:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在另外的實施例中,一種或多種分化標記物基因選自T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。
在其他實施例中,減少的自發分化維持至少5代。
在某些實施例中,減少的自發分化維持至少10代。
在另外的實施例中,減少的自發分化維持至少50代。
在特定實施例中,減少的自發分化維持至少100代。
在進一步的實施例中,前述方法包括在傳代期間解離一個或多個iPSC。
在特定實施例中,一個或多個iPSC的活力在傳代期間得到維持。
在其他實施例中,一個或多個iPSC包含正常核型。
在另外的實施例中,一個或多個iPSC在無飼養者環境中進行培養。
在某些實施例中,一個或多個iPSC的基因組穩定性維持至少10代。
在另外的實施例中,一個或多個iPSC的基因組穩定性維持至少50代。
在特定實施例中,一個或多個iPSC的基因組穩定性維持至少100代。
在各個特定實施例中,本發明部分提供了根據前述實施例中任何一個產生的包含基態多能性的誘導多能幹細胞(iPSC)。
在各個實施例中,本發明部分提供了包含基態多能性的誘導多能幹細胞(iPSC),其中所述iPSC不包含編碼重編程因子多肽的外源引入的多核苷酸。
在一些實施例中,本發明提供了用於使非多能細胞重編程為多能細胞的方法,所述方法包括將下述引入非多能細胞內:(i)編碼選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少一種多肽的一種或多種多核苷酸,或(ii)選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少一種多肽,由此使非多能細胞重編程為多能細胞。
在一個特定實施例中,引入包括(i)引入編碼OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的一種或多種多核苷酸,或(ii)引入OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽。
在另一個實施例中,引入包括(i)引入編碼OCT4多肽、ECAT1多肽和UTF1多肽的一種或多種多核苷酸,或(ii)引入OCT-4多肽、ECAT1多肽和UTF1多肽。
在一些實施例中,一種或多種多核苷酸通過逆轉錄病毒、仙臺病毒、腺病毒、附加體、微環、具有表達盒的載體系統或mRNA而引入。
在一個特定實施例中,逆轉錄病毒是慢病毒。
在另一個實施例中,多能細胞不含外源多核苷酸。
在另一個實施例中,一種或多種多核苷酸通過CRE介導的切除進行切除。
在一些實施例中,該方法還包括將下述引入非多能細胞內:(i)編碼SV40LT多肽的多核苷酸,或(ii)SV40LT多肽。
在一個實施例中,該方法還包括使所述非多能細胞與下述中的至少一種接觸:TGFβR抑制劑、Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑,其中所述Wnt途徑激動劑任選是GSK3抑制劑。
在另一個實施例中,非多能細胞與下述接觸:TGFβR抑制劑、Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑,其中所述Wnt途徑激動劑任選是GSK3抑制劑。
在一個特定實施例中,該方法還包括在包含Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑的培養基中培養多能細胞,其中所述Wnt途徑激動劑任選是GSK3抑制劑,並且其中所述培養基不包含TGFβR抑制劑。
在一些實施例中,ROCK抑制劑是thiazovivin或Y27632,TGFβR抑制劑是A-83-01或SB431542,GSK3抑制劑是CHIR99021或BIO,或MEK抑制劑是PD98059或PD032901。
在另一個實施例中,多能細胞的多能性維持至少5次細胞分裂或至少10次細胞分裂。
在一個特定實施例中,一種或多種多核苷酸作為包含多重多核苷酸的多順反子載體引入,所述多重多核苷酸通過至少一個2A肽分開。
在一個實施例中,多順反子載體包含編碼OCT4多肽的多重多核苷酸。
在一些實施例中,該方法包括通過在多能細胞中選擇OCT4表達來鑑定多能細胞。
在一個實施例中,OCT4表達的選擇包括異位Oct-4表達的選擇。
在另一個實施例中,培養產生多能幹細胞群。
在另一個實施例中,多能幹細胞群是至少70%同質的、至少80%同質的、或至少90%同質的。
在另外一個實施例中,多能細胞群的至少70%、至少80%、或至少90%表達SSEA和Tra-181。
在一個實施例中,多能細胞或多能細胞群能夠單細胞傳代。
在一個實施例中,通過單細胞傳代產生的細胞具有正常核型。
在一個實施例中,本發明提供了根據上文方法中任何一種的方法產生的多能細胞。
在另一個實施例中,本發明提供了包含經分離的非多能細胞的組合物,所述經分離的非多能細胞包含(i)編碼選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少一種多肽的一種或多種外源多核苷酸,或(ii)選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少一種外源多肽。
在一個實施例中,細胞包含(i)編碼OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽中的至少兩種的一種或多種外源多核苷酸,或(ii)選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少兩種外源多肽。
在另一個實施例中,細胞包含(i)編碼OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽中的至少三種的一種或多種外源多核苷酸,或(ii)選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少三種外源多肽。
在一個特定實施例中,細胞包含(i)編碼OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的一種或多種外源多核苷酸,或(ii)外源OCT4多肽、外源ECAT1多肽、外源UTF1多肽、外源NANOG多肽和外源ESRRB多肽。
在另一個實施例中,細胞包含編碼OCT4多肽、ECAT1多肽和UTF1多肽的一種或多種外源多核苷酸,或外源OCT4多肽、外源ECAT1多肽和外源UTF1多肽。
在一個實施例中,已使細胞與TGFβR抑制劑、GSK3抑制劑、MEK抑制劑和ROCK抑制劑中的至少一種接觸。
在另一個實施例中,已使細胞與TGFβR抑制劑、Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑接觸,其中所述Wnt途徑激動劑任選是GSK3抑制劑。
在一個特定實施例中,ROCK抑制劑是thiazovivin或Y27632,TGFβR抑制劑是A-83-01或SB431542,GSK3抑制劑是CHIR99021或BIO,或MEK抑制劑是PD98059或PD032901。
在另一個實施例中,一種或多種外源多核苷酸通過逆轉錄病毒、仙臺病毒、腺病毒、附加體、微環、具有表達盒的載體系統或mRNA而引入非多能細胞。
在另一個實施例中,逆轉錄病毒是慢病毒。
在一個特定實施例中,細胞不含外源多核苷酸。
在一個實施例中,一種或多種外源多核苷酸通過CRE介導的切除進行去除。
在另一個實施例中,細胞包含編碼SV40LT抗原多肽的外源多核苷酸,或外源SV40LT抗原多肽。
在一個實施例中,一種或多種外源多核苷酸作為包含多重多核苷酸的多順反子載體引入,所述多重多核苷酸通過至少一個2A肽分開。
在另一個實施例中,多順反子載體包含編碼OCT4多肽的多重多核苷酸。
在一個實施例中,編碼OCT4多肽的外源多核苷酸連接至可選標記物。
在一個特定實施例中,本發明提供了由下述中的至少一種、至少兩種或至少三種組成的組合物:(i)編碼OCT4多肽的cDNA、(ii)編碼ECAT1多肽的cDNA、(iii)編碼UTF1多肽的cDNA、(iv)編碼NANOG多肽的cDNA、和(v)編碼ESRRB多肽的cDNA。
在一個實施例中,組合物由編碼OCT4多肽的cDNA、編碼ECAT1多肽的cDNA、編碼UTF1多肽的cDNA、編碼NANOG多肽的cDNA、和編碼ESRRB多肽的cDNA組成。
在另一個實施例中,組合物由編碼OCT4多肽的cDNA、編碼ECAT1多肽的cDNA和編碼UTF1多肽的cDNA組成。
在一個特定實施例中,每種cDNA在逆轉錄病毒、仙臺病毒、腺病毒、附加體、微環、具有表達盒的載體系統或mRNA中編碼。
在一個實施例中,本發明提供了包含一種或多種多核苷酸的載體,所述一種或多種多核苷酸編碼選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽的至少一種重編程因子多肽。
在一個實施例中,一種或多種多核苷酸編碼OCT多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽。
在另一個實施例中,一種或多種多核苷酸編碼OCT多肽、ECAT1多肽和UTF1多肽。
在另一個特定實施例中,載體還包含編碼SV40LT抗原多肽的多核苷酸。
在一個實施例中,載體是逆轉錄病毒、仙臺病毒、腺病毒、附加體、微環、具有表達盒的載體系統或mRNA。
在一個實施例中,逆轉錄病毒是慢病毒。
在一個特定實施例中,本發明提供了用於使非多能細胞重編程為多能細胞的試劑盒,所述試劑盒包括:編碼選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽或ESRRB多肽的至少一種多肽的一種或多種多核苷酸;或選自OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽或ESRRB多肽的至少一種多肽;和TGFβR抑制劑、Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑中的至少一種,其中所述Wnt途徑激動劑任選是GSK3抑制劑。
在一個實施例中,一種或多種多核苷酸編碼OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽,或至少一種多肽包含OCT4多肽、ECAT1多肽、UTF1多肽、NANOG多肽和ESRRB多肽。
在另一個實施例中,一種或多種多核苷酸編碼OCT4多肽、ECAT1多肽和UTF1多肽,或至少一種多肽包含OCT4多肽、ECAT1多肽和UTF1多肽。
在一個實施例中,試劑盒包括編碼SV40LT抗原多肽的多核苷酸,或SV40LT抗原多肽。
在另一個實施例中,試劑盒包括TGFβR抑制劑、Wnt途徑激動劑、MEK抑制劑和ROCK抑制劑,其中所述Wnt途徑激動劑任選是GSK3抑制劑。
在一個實施例中,ROCK抑制劑是thiazovivin或Y27632,TGFβR抑制劑是A-83-01或SB431542,GSK3抑制劑是CHIR99021或BIO,或MEK抑制劑是PD98059或PD032901。
在另一個實施例中,至少一種多核苷酸在逆轉錄病毒、仙臺病毒、腺病毒、附加體、微環、具有表達盒的載體系統或mRNA中編碼。
在另一個實施例中,逆轉錄病毒是慢病毒。
在一個實施例中,至少一種多核苷酸在多順反子載體中編碼,其中每種多核苷酸通過2A肽分開。
在另外一個實施例中,多順反子載體包含編碼OCT4多肽的兩種或更多種多核苷酸。
在一個特定實施例中,編碼OCT4多肽的至少一種多核苷酸連接至可選標記物。
在另一個特定實施例中,本發明提供了用於產生多能幹細胞群的方法,所述方法包括:提供非多能細胞群;將連接至可選標記物的編碼OCT4多肽的多核苷酸引入非多能細胞群內;在足以使非多能細胞群的至少一部分重編程為多能細胞的條件下,使非多能細胞群與多核苷酸一起溫育;選擇表達可選標記物的細胞,由此提供多能幹細胞群。
在另一個實施例中,多核苷酸作為多順反子載體引入,所述多順反子載體包含編碼OCT4多肽的多重多核苷酸。
在一個實施例中,多重多核苷酸通過至少一個2A肽分開。
在另外一個實施例中,細胞群中的至少10%的細胞表達SSEA和TRA-181。
附圖說明
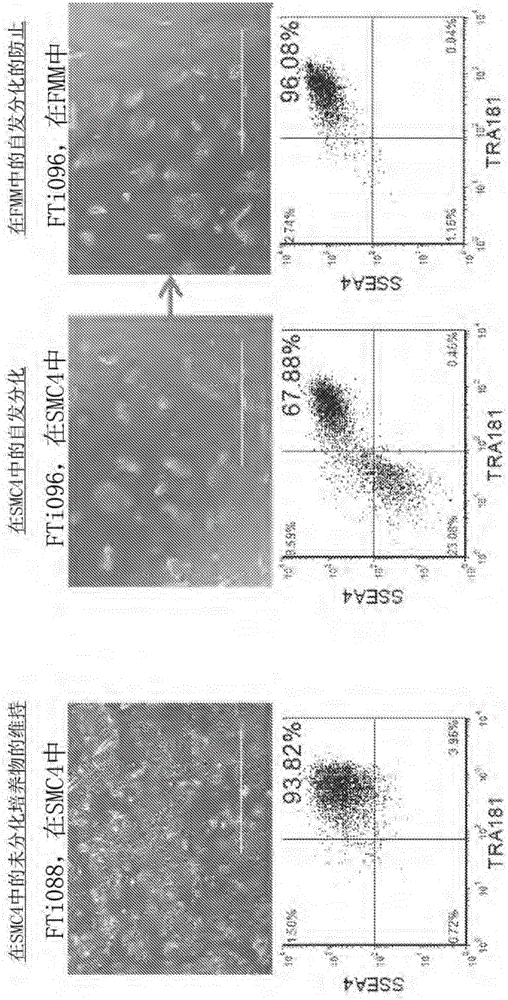
圖1A-1E顯示了來自關於增強的重編程和hiPSC維持的多階段培養平臺的結果。(1A)慢病毒生成的hiPSC克隆FTi088在SMC4中維持非分化細胞的同質群,而自發分化在SMC4中培養的慢病毒生成的hiPSC系FTi096中可見。當FTi096轉變為FMM共3次傳代時,自發分化降到最低,如通過形態(上圖)以及關於SSEA4和TRA1-81的流式細胞術(下圖)所示。(1B)關於病毒元件WPRE的轉基因表達的qRT-PCR。表達針對GAPDH且相對於慢病毒感染後四天(第4天P.I.)親本成纖維細胞系的WPRE表達進行標準化。未感染的成纖維細胞系(成纖維細胞)和人ESC系HUES9用作陰性對照。每個集合的值在條上方指出。(1C)在10次傳代後,各種培養基組分對無轉基因慢病毒誘導的hiPSC群的SSEA4和TRA1-81的作用的篩選;SB431542(-TGFβRi)的去除從10增加到100ng/mL bFGF,添加10ng/mL LIF。(1D)成纖維細胞細胞系用含有基因集合OCT4/KLF4/SOX2的慢病毒構建體進行轉染,並且拆分到各種培養基(常規培養基;Conv.)內,培養17天且在第17天時就SSEA4和TRA1-81雙陽性群進行分選。分選門以藍色突出顯示。每個集合在分別的培養基中培養另外10天,除了拆分到FMM和SMC4內的SMC4集合之外。在第27天時,培養物就SSEA4和TRA1-81雙陽性群進行重新分選,以分選事件的標準化密度種植且在分別的培養基中維持另外9天。將常規培養集合門控擴大,以實現標準化數目的細胞。在第36天時,每種培養物就OCT4和NANOG表達進行染色。關於每個集合的代表性免疫細胞化學圖像顯示於右圖中。(1E)第36天染色的集落計數如(1D)中討論的。誤差條代表關於FRM至FMM和FRM的一式三份以及關於FMM和hESC的一式兩份。
圖2A-2E顯示了個別附加體重編程的hiPSC被有效選擇且種植在96孔板中用於克隆擴增。(2A)附加體誘導、多階段培養平臺、流式細胞術分選和克隆擴增的示意性時機圖示。(2B)在所示的轉染後天數時,在FF培養(在2A中概述)中的FRM至FMM轉變中維持的附加體誘導的重編程的流式細胞術概況。用於每種親本系(SSEA4+/TRA1-81+/CD30+群)的分選門控策略以分別的顏色示出,對應於代表含有個別hiPSC克隆的96孔板的孔百分比的底部直方圖小圖。含有多重克隆或分化克隆的孔不進行評分。實線代表在所有衍生中的平均百分比,而虛線代表標準差。(2C)在MEF細胞的存在下,在常規培養基中維持的轉染後19天誘導至重編程的FTC007的流動概況。誘導群取自(2B)中的相同FTC007群,然而,其後在不同培養中進行處理。(2D)在96孔板中的分選集落的各種多能性標記物的免疫細胞化學分析。右下角小圖代表DAPI染色。(2E)關於以3個細胞/孔的SSEA4/TRA1-81/CD30直接分選的(FACS)96孔板的每個孔的NANOG表達的qRT-PCR。表達範圍在如圖例中所述相對於H1人ESC且針對GAPDH標準化的零至四倍表達之間。
圖3A-3F顯示了附加體重編程的hiPSC克隆維持其未分化狀態且不含轉基因序列。(3A)在單細胞傳代後24小時,hiPSC克隆的典型形態。(3B)在培養期間hiPSC克隆的代表性圖像。(3C)關於來源於各種hiPSC克隆的附加型DNA的PCR分析。泳道1,FTC007-c1p4;泳道2,FTC007-c21p4;泳道3,FTC016-c25p5;泳道4,FTC016-c36p5;泳道5,FTC017-c11p7;泳道6,FTC017-c14p7;泳道7,FTC017-c17p6(用作陽性對照的維持附加體構建體的系);泳道8,未經轉染的FTC007;泳道9,使用慢病毒構建體生成的hiPSC(以充當針對交叉感染的對照);泳道10,用作陽性對照的附加型載體。100ng基因組DNA的輸入和35個PCR循環用於所有集合。(3D)對於OCT4、NANOG、TRA1-81和TRA160的表達,通過免疫螢光檢測的多能性標記物。(3E)關於來自各種親本系的所選擇的hiPSC克隆的流式細胞術概況。上面一行概括SSEA4/TRA1-81表面表達。下面一行概括OCT4/NANOG細胞內表達。(3F)關於內源多能基因表達的qRT-PCR分析。數據針對GAPDH且相對於HUES9hESC進行標準化。在KLF4表達的情況下,兩個數據點比HUES9大超過15倍且在圖上註明。誤差條代表重複的標準差。
圖4A-4E顯示了基因組穩定性和多能性在連續單細胞和FF培養中得到維持。(4A)來自在FF和單細胞培養中維持的各種hiPSC克隆的20至40個G帶中期細胞的細胞遺傳學分析。(4B)在FF和單細胞培養中長期傳代(p25-30)的hiPSC克隆的流式細胞術概況和細胞遺傳學分析。(4C)FTC017-c11三至四天的定向分化。(4D)hiPSC克隆的胚狀體形成和分化證實三系分化。免疫細胞化學在分化後28天進行:外胚層,TUJ1;中胚層,α平滑肌肌動蛋白(aSMA);內胚層,AFP。(4E)來源於代表每個體細胞譜系的FTC007-c21和FTC016-c25的畸胎瘤的組織切片。黑色箭頭,內胚層;白色箭頭,外胚層;灰色箭頭,中胚層。
圖5A-5J顯示了具有最低限度數目的重編程因子的hiPSC克隆的衍生。(5A)將OCT4、SOX2和NANOG以各種形式克隆到pCEP4內。表代表載體系統和縮寫。(5B)在誘導後第13天時,通過各種基因組合誘導的重編程動力學的SSEA4和TRA1-81流式細胞術概況。(5C)代表以3和9個細胞/孔在96孔板的孔中TRA1-81陽性hiPSC克隆的存在的效率直方圖。(5D)關於來源於各種hiPSC克隆的附加型DNA的PCR分析。泳道1 2xO+OS+ONS+T-c7p6;泳道2,2xO+OS+ONS+T-c10p6;泳道3,2xO+ONS+T-c5p5;泳道4,2xO+ONS+T-c9p5;泳道5,2xO+OS+T-c7p7;泳道6,2xO+OS+T-c9p6;泳道7,未經轉染的FTC007;泳道8,使用慢病毒構建體生成的hiPSC;泳道9,用作陽性對照的附加型載體。100ng基因組DNA的輸入和35個PCR循環用於所有集合。(5E)來源於2xO+OS+T的克隆9的形態。(5F)關於OCT4、NANOG、TRA1-81和TRA160的表達,通過免疫螢光檢測的多能性標記物。圖像以10x放大率獲得。(5G)來源於所選擇的基因集合的hiPSC克隆的流動概況。上面一行概括SSEA4/TRA1-81表面表達。下面一行概括OCT4/NANOG細胞內表達。(5H)轉染後大約72至96小時,所選擇的hiPSC克隆的定向分化。(5I)來自在FF和單細胞培養中維持的各種hiPSC克隆的G帶中期細胞的細胞遺傳學分析。(5J)來源於代表每個體細胞譜系的hiPSC克隆2xO+OS+ONS+T-c10的畸胎瘤的組織切片。左圖,內胚層;中圖,中胚層;右圖,外胚層。
圖6A-6B顯示了在FMM中最低限度因子附加體誘導的hiPSC的相對基因表達概況。熱圖結果來源於Fluidigm動態陣列,描述常規維持的hiPSC系、常規維持的H1hESC以及在FMM中維持的使用各種基因組合衍生的附加型hiPSC系的多能性(6A)和分化(6B)基因的相對基因表達水平(RQ)。關於每個系的相對基因表達在每個框內註明,並且基於三個表達水平編碼的顏色概括於圖例中(右下)。所有集合一式兩份進行,針對兩種持家基因(GAPDH和HPRT1)的平均表達進行標準化,並且參考代表1x值的六個對照常規系(在MEF上的OSK hiPSC和H1hESC)的中值表達水平。
圖7A-7G顯示了FMM維持的hiPSC具有分化基因的減少表達且代表基態。(7A)總共339個探針組在常規和FMM培養之間差異表達大於或小於2.5倍。基於歐幾裡得距離測量,使用完全連鎖方法,關於339個探針組的分層聚類。(7B)對於常規培養(與FMM培養相比較),213個探針組的基因本體生物過程富集分析(D.A.V.I.D.)上調2.5倍或更大。(7C)代表基態或亞穩多能性態的基因列表。列表來源於正文中註明的參考文獻。7(D)基於歐幾裡得距離測量,使用完全連鎖方法,關於對應於(7C)中的基因的231個探針組的分層聚類。(7E)關於對應於(7C)中的基因的探針組的RMA(log2)強度。左圖代表關於基態的39個探針組,右圖代表關於亞穩態的188個探針組。平均常規培養強度水平在X軸上標繪,而平均FMM/SMC4強度在Y軸上標繪,黑線指示相等表達。(7F)使用Affymetrix探針組,在衍生且在常規培養基培養中培養的hiPSC克隆和其適應SMC4培養的配對物之間的X染色體定位基因的基因表達比較。與XIST基因表達相關的探針組是突出顯示的。(7G)在FMM中維持或適應常規培養共5代的hiPSC克隆上的HEK27me3的代表性圖像。左圖中的虛線箭頭指向不存在H3K27me3染色的代表性核,而右圖中的實線箭頭指向對於H3K27me3染色陽性的核。核陽性染色百分比在每個小圖的左下側指出。FMM培養的細胞具有更大的核。比例尺=50μm。
圖8A-8D顯示了對於FRM和FMM附加體誘導的重編程。(8A)重編程庫的第10天SSEA4和TRA1-81流動概況。(8B)在重編程期間可見的典型集落的代表性形態。圖像在轉染後第13天時獲取。(8C)對於前14天在FRM中維持的附加體重編程成纖維細胞進行拆分,且在FRM中維持或轉變成FMM。重編程培養物隨後在轉染後第21天時就SSEA4/TRA1-81/CD30進行分選,並且在分析前在FRM或FMM中維持另外10天。(8D)在FRM或FMM中的代表性培養物的形態和流動概況。白色箭頭指向培養物中的分化細胞區域,所述培養物由未分化和分化群的混合物組成。黑色箭頭指向大部分未分化群的尖銳邊緣。下圖是代表性流動概況。FSC;前向側散射。
圖9A-9B顯示了各種親本系的重編程。(9A)在該研究中使用的起始細胞系的概括表。除與每個系相關的具體信息之外,註明了在附加體轉染後分選時的陽性SSEA4/TRA1-81/CD30群百分比。(9B)描述CD34富集的臍帶血細胞的分選和培養的圖示。先前在庫中維持的0.5ml體積的臍帶血用於提取65,000個CD34+CD45+Lin-細胞,所述細胞在附加體轉染之前懸浮培養6天。
圖10A-10G顯示了在重編程和維持過程期間hiPSC的表徵。(10A)單細胞96孔板分選後三天的典型集落形態。比例尺代表400μm。(10B)由各種起始細胞分選後7-9天,單細胞衍生的hiPSC樣集落的代表性形態。比例尺代表1000μm。(10C)關於在96孔板中的hiPSC樣集落的NANOG表達的免疫細胞化學。(10D)誘導為重編程且在Matrigel或玻連蛋白塗布的培養板上維持的FTC007的第16天流動概況分析。(10E)亮視野圖像,(10F)關於OCT4和NANOG的免疫螢光,或(10G)關於在Matrigel上連續或關於5次傳代玻連蛋白的在FMM中維持的FTC016-c28的SSEA4和TRA1-81的流式細胞術分析。
圖11A-11C顯示了對於FMM培養平臺,最低限度基因重編程的例子。(11A)使用含有潮黴素選擇盒的附加型構建體,從轉染後第2天到第5天用潮黴素處理的細胞的形態。(11B)重編程庫維持更長持續時間且在轉染後第16天時進行概況分析。(11C)在Matrigel或玻連蛋白上維持的培養物外觀。
圖12A-12E顯示了在多重條件下培養的hiPSC的特徵。(12A)慢病毒衍生和SMC4維持的FTi111展示多能性的標誌且維持基因組完整性。(12B)FTi111p43的解凍策略描述。單個小瓶如註明的解凍到四個培養環境內。存活培養物在分別培養中進行傳代,除了在飼養細胞上補充有thiazovivin的常規培養之外,所述常規培養在飼養細胞的存在下轉變為不含Thiazovivin的常規培養,且作為塊進行傳代。(12C)在解凍後的各種培養中的回收細胞的形態。在飼養細胞的存在下,在不含Thiazovivin的常規培養中未鑑定到存活細胞。(12D)在解凍後的第3代時的培養集合的形態。更大的集落形態與常規培養相關。比例尺1000μm。(12E)關於每個培養集合的內源多能基因表達的qRT-PCR分析。數據針對GAPDH且相對於H1hESC進行標準化。
圖13A-13E顯示了在各種條件下培養的hiPSC的基因表達概況的基因本體學。(13A)描述在總體基因表達研究中所述的每個系的衍生和維持的表。(13B)總共300個探針組在常規和小分子(FMM和SMC4)培養條件之間差異表達大於或小於2.5倍。基於歐幾裡得距離測量,使用完全連鎖方法,關於300個探針組的分層聚類。(13C)對於常規培養(與小分子培養相比較),133個探針組的基因本體生物過程富集分析(D.A.V.I.D.)上調2.5倍或更大。(13D)對於小分子培養(與常規培養相比較),167個探針組的基因本體生物過程富集分析(D.A.V.I.D.)上調2.5倍或更大。(13E)對於FMM培養(與常規培養相比較),126個探針組的基因本體生物過程富集分析上調2.5倍或更大。
圖14A-14F顯示了示出用於重編程的慢病毒構建體(14A-14B)和附加型構建體(14C-14F)的例子的克隆圖。慢病毒構建體包括EF1α啟動子和關於CRE介導的轉基因切除的LOXP位點。附加型構建體還包括EF1α啟動子。
圖15A-15C顯示了在第8-15天時,關於各種重編程因子組合的代表性流動分析。人成纖維細胞用慢病毒介導的重編程因子包括OCT4、ECAT1和UTF1的各種組合進行誘導。
圖16A-16D顯示了在第21-27天時,關於各種重編程因子組合的代表性流動分析和iPSC形態特徵。數據顯示獨特重編程組合可用於衍生SSEA4+/TRA181+hiPSC。
圖17A-17B顯示了如通過流動分析(SSEA4+/TRA181+和CD30+群)和iPSC形態特徵示出的高度增強的慢病毒重編程效率。人成纖維細胞用OCT4、ECAT1、UTF1、ESRRB和NANOG進行重編程。細胞使用FRM進行重編程且在FMM中維持。
圖18A-18D顯示了在96孔分選後,在7-9次傳代後,建立的四個iPSC克隆的代表性流動分析和相圖像。克隆用慢病毒重編程因子(OCT4、ECAT1、UTF1、ESRRB和NANOG)與FRM生成且在FMM中維持。表達高SSEA4+/TRA181+的群指示多能性。
圖19A-19B顯示了在用慢病毒重編程因子OCT4、ECAT1、UTF1、NANOG和ESRRB重編程的人成纖維細胞中的OCT4和NANOG表達的代表性流動分析。克隆使用FRM進行重編程且在FMM中維持。表達高OCT4+/NANOG+的群指示多能性。
圖20A-20C顯示了來源於用慢病毒重編程因子OCT4、ECAT1、UTF1、NANOG和ESRRB重編程的人成纖維細胞的hIPSC克隆的核型分析。克隆使用FRM進行重編程且在FMM中維持。克隆顯示出正常、雄性核型。
圖21顯示了與重編程因子組合OCT4/NANOG/SOX2/LARGE T相比較,重編程因子組合OCT4/ESRRB/NANOG/ECAT1/UTF1的96孔板分選效率。
圖22A-22B顯示了(22A)在4、6和11天時,用OCT4-P2A-OCT4/NANOG-P2A-ESRRB-T2A-LIN28/ECAT1-T2A-UTF1,和(22B)對於兩個孔在7天時和一個孔在10天時,用OCT4-P2A-ESRRB/OCT4-P2A-NANOG/ECAT1-T2A-UTF1重編程的細胞的96孔擴增期間的集落圖像。
圖23A-23C顯示了(23A)證實重編程因子化學計量學的效應的流動分析和使用遺傳標記物用於選擇具有異位OCT4表達的細胞的結果概括,(23B)關於用附加型OCT4-P2A-NANOG-T2A-SOX2/SV40大T抗原重編程,不含OCT4的選擇的人成纖維細胞的流動分析,和(23C)關於用附加型OCT4-P2A-NANOG-T2A-SOX2/SV40大T抗原/OCT2-P2A-OCT4-嘌呤黴素重編程的人成纖維細胞的流動分析。
圖24顯示了對於多能性標記物OCT4(綠色)和TRA181(紅色)用以藍色的DAPI染色的衍生的iPSC克隆的免疫螢光分析。
圖25顯示了CRE介導的切除後,SSEA4+/TRA181+/CD30+96孔板分選的克隆的圖像。集落由最初來源於人成纖維細胞的iPSC克隆進行分選,用慢病毒因子OCT4、ECAT1、UTF1、NANOG和ESRRB進行重編程,並且隨後就轉基因進行切除。分選的集落顯示iPSC表型。
具體實施方式
A.概述
用於多能細胞的生產和維持的現有方法仍未實現無足跡多能細胞的同質培養,所述多能細胞不含自發分化且順應高解析度/高克隆形成能力單細胞傳代和大規模擴增。基態多能細胞可賦予克服這些挑戰的質量和特徵。然而,迄今為止,不存在在無飼養者條件下基態多能細胞的高流通量生成的可靠或穩健的方法。因此,現有方法可能不適合於生產工業或臨床級別的多能細胞。本文考慮的本發明解決了在基態多能性中或具有基態多能性特徵的穩定多能細胞的穩健生成的需要,並且解決了在適合於工業和臨床使用的穩定多能細胞製造中的問題。
一般而言,本發明涉及用於多能細胞,特別是具有減少的自發分化的細胞,包括基態多能細胞的改良製造的組合物和方法。更特別地,本發明涉及以階段特異性方式,利用細胞信號轉導途徑的小分子調製劑的多階段培養平臺,且使多能細胞能夠衍生和維持至其中培養方法和衍生多能細胞的方法不再是下遊使用的變化性和/或門控活性源的點。此外,本文考慮的培養平臺使多能細胞在無飼養者條件下的衍生和維持成為可能,具有改善的基因組穩定性、改善的未分化狀態、減少的自發分化、改善的培養同質性、改善的培養中的存活、解離和單個多能細胞的傳代,以及使細胞無轉基因或足跡重編程為基態多能性的改良方法。因此,本文考慮的組合物和方法使多能細胞和/或基態多能細胞的製造成為可能,所述多能細胞適合於工業和臨床用途。
迄今為止,小分子驅動的平臺未證實增強來源於人細胞的無足跡誘導多能幹細胞(iPSC)的重編程且支持單細胞和FF培養的能力(Nichols和Smith,2012)。本文考慮的培養平臺部分提供了小分子抑制劑的特定組合以階段特異性方式的應用,以使多能幹細胞能夠快速和穩健的重編程以及穩定的長期培養。在各個實施例中,本發明提供了用於誘導或維持改善的未分化多能態包括基態多能性的培養平臺。本文考慮的平臺還提供了用於生產和維持人iPSC(hiPSC)中的基態多能性的穩健培養系統。在一個實施例中,培養平臺使無轉基因或足跡的重編程方法成為可能。在特定實施例中,本文考慮的平臺代表用於製造hiPSC的改良方法,所述方法克服在無轉基因hiPSC的多路衍生和維持中的關鍵挑戰。
除非特別指出相反,否則本發明的實踐將採用在本領域技術內的化學、生物化學、有機化學、分子生物學、微生物學、重組DNA技術、遺傳學、免疫學、細胞生物學、幹細胞方案、細胞培養和轉基因生物學的常規方法,其中許多在下文描述用於舉例說明的目的。此類技術在文獻中充分說明。參見例如Sambrook等人,Molecular Cloning:A Laboratory Manual(第3版,2001);Sambrook等人,Molecular Cloning:A Laboratory Manual(第2版,1989);Maniatis等人,Molecular Cloning:A Laboratory Manual(1982);Ausubel等人,Current Protocols in Molecular Biology(John Wiley and Sons,2008年7月更新);Short Protocols in Molecular Biology:A Compendium of Methods from Current Protocols in Molecular Biology,Greene Pub.Associates and Wiley-Interscience;Glover,DNA Cloning:A Practical Approach,第I&II卷(IRL Press,Oxford,1985);Anand,Techniques for the Analysis of Complex Genomes,(Academic Press,New York,1992);Guthrie和Fink,Guide to Yeast Genetics and Molecular Biology(Academic Press,New York,1991);Oligonucleotide Synthesis(N.Gait編輯,1984);Nucleic Acid Hybridization(B.Hames&S.Higgins編輯,1985);Transcription and Translation(B.Hames&S.Higgins編輯,1984);Animal Cell Culture(R.Freshney編輯,1986);Perbal,A Practical Guide to Molecular Cloning(1984);Fire等人,RNA Interference Technology:From Basic Science to Drug Development(Cambridge University Press,Cambridge,2005);Schepers,RNA Interference in Practice(Wiley-VCH,2005);Engelke,RNA Interference(RNAi):The Nuts&Bolts of siRNA Technology(DNA Press,2003);Gott,RNA Interference,Editing,and Modification:Methods and Protocols(Methods in Molecular Biology;Human Press,Totowa,NJ,2004);Sohail,Gene Silencing by RNA Interference:Technology and Application(CRC,2004);Clarke和Sanseau,microRNA:Biology,Function&Expression(Nuts&Bolts series;DNA Press,2006);Immobilized Cells And Enzymes(IRL Press,1986);the treatise,Methods In Enzymology(Academic Press,Inc.,N.Y.);Gene Transfer Vectors For Mammalian Cells(J.H.Miller和M.P.Calos編輯,1987,Cold Spring Harbor Laboratory);Harlow和Lane,Antibodies,(Cold Spring Harbor Laboratory Press,Cold Spring Harbor,N.Y.,1998);Immunochemical Methods In Cell And Molecular Biology(Mayer和Walker編輯,Academic Press,London,1987);Handbook Of Experimental Immunology,第I-IV卷(D.M.Weir和C.Blackwell編輯,1986);Riott,Essential Immunology,第6版,(Blackwell Scientific Publications,Oxford,1988);Embryonic Stem Cells:Methods and Protocols(Methods in Molecular Biology)(Kurstad Turksen編輯2002);Embryonic Stem Cell Protocols:Volume I:Isolation and Characterization(Methods in Molecular Biology)(Kurstad Turksen編輯,2006);Embryonic Stem Cell Protocols:Volume II:Differentiation Models(Methods in Molecular Biology)(Kurstad Turksen編輯,2006);Human Embryonic Stem Cell Protocols(Methods in Molecular Biology)(Kursad Turksen編輯,2006);Mesenchymal Stem Cells:Methods and Protocols(Methods in Molecular Biology)(Darwin J.Prockop,Donald G.Phinney和Bruce A.Bunnell編輯,2008);Hematopoietic Stem Cell Protocols(Methods in Molecular Medicine)(Christopher A.Klug和Craig T.Jordan編輯,2001);Hematopoietic Stem Cell Protocols(Methods in Molecular Biology)(Kevin D.Bunting編輯,2008)Neural Stem Cells:Methods and Protocols(Methods in Molecular Biology)(Leslie P.Weiner編輯2008);Hogan等人,Methods of Manipulating the Mouse Embyro(第2版,1994);Nagy等人,Methods of Manipulating the Mouse Embryo(第3版,2002),和The Zebrafish book.A guide for the laboratory use of zebrafish(Danio rerio),第4版,(Univ.of Oregon Press,Eugene,OR,2000)。
本文引用的所有出版物、專利和專利申請在此以引用的方式全文併入。
B.定義
除非另有定義,否則本文使用的所有技術和科學術語具有與本發明所屬領域普通技術人員通常理解相同的含義。為了本發明的目的,下文定義了下述術語。
冠詞「一個」、「一種」和「該/所述」在本文中用於指一個或超過一個(即至少一個)冠詞的語法對象。例如,「元件」意指一個元件或超過一個元件。
替代(例如「或」)的使用應理解為意指替代方案中任一、兩者或其任何組合。
術語「和/或」應理解為意指替代方案中任一或兩者。
如本文使用的,術語「約」或「大約」指與參考數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度相比較,改變多達15%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度。在一個實施例中,術語「約」或「大約」指圍繞參考數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度±15%、±10%、±9%、±8%、±7%、±6%、±5%、±4%、±3%、±2%或±1%的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度範圍。
如本文使用的,術語「基本上」或「大體上」指與參考數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度相比較,約90%、91%、92%、93%、94%、95%、96%、97%、98%或99%或更高的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度。在一個實施例中,術語「大體上相同」或「基本上相同」指與參考數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度大約相同的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度範圍。
如本文使用的,術語「基本上不含」和「大體上不含」可互換使用,並且當用於描述組合物例如細胞群或培養基時,指不含指定物質,例如95%不含、96%不含、97%不含、98%不含、99%不含指定物質的組合物,或如通過常規手段測量是無法檢測的。類似含義可應用於術語「不存在」,當指不存在組合物的特定物質或組分時。
如本文使用的,術語「可觀的」指通過一種或多種標準方法可容易檢測的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度範圍。術語「不可觀的(not-appreciable)」和「不可觀的(not appreciable)」以及等價物指通過標準方法無法容易檢測或無法檢測的數量、水平、值、數目、頻率、百分比、尺度、大小、量、重量或長度範圍。在一個實施例中,如果事件小於5%、4%、3%、2%、1%、0.1%、0.01%、0.001%或更少的時間發生,則它是不可觀的。
本說明書自始至終,除非上下文另有要求,否則單詞「包含(comprise)」、「包含(comprises)」和「包含(comprising)」應理解為暗示包括所述步驟或元件或者步驟或元件組,但不排除任何其他步驟或元件或者步驟或元件組。在特定實施例中,術語「包括」、「具有」、「含有」和「包含」同義使用。
「由……組成」意指包括但限制於在短語「由……組成」後的任何。因此,短語「由……組成」指示所列出的元件是必需或強制性的,並且可不存在其他元件。
「基本上由……組成」意指包括在短語後列出的任何元件,並且限制於不幹擾或促成所列出的元件的公開內容中指定的活性或動作的其他元件。因此,短語「基本上由……組成」指示所列出的元件是必需或強制性的,但其他元件並非任選的,並且取決於它們是否影響所列出的元件的活性或動作,可存在或不存在。
本說明書自始至終提及「一個實施例」、「實施例」、「特定實施例」、「相關實施例」、「某個實施例」、「另外的實施例」或「進一步的實施例」或其組合意指與實施例結合描述的特定特點、結構或特徵包括在本發明的至少一個實施例中。因此,前述短語在本說明書自始至終的各個地方的出現不一定全部指相同實施例。此外,特定特點、結構或特徵可以任何合適方式在一個或多個實施例中組合。
術語「離體」一般指在生物外發生的活性,例如在生物外的人工環境中在活組織中或活組織上完成的實驗或測量,優選伴隨天然條件的最低限度改變。在特定實施例中,「離體」程序涉及從生物中獲得的活細胞或組織,並且在實驗室儀器中培養,通常在無菌條件下,且通常為數小時或最高達約24小時,但包括最高達48或72小時,取決於環境。在某些實施例中,可收集且冷凍此類組織或細胞,並且隨後解凍用於離體處理。使用活細胞或組織持續長於幾天的組織培養實驗或程序通常視為「在體外」,儘管在某些實施例中,該術語可與離體互換使用。
術語「在體內」一般指在生物內發生的活性。
如本文使用的,術語「重編程」或「去分化」或「增加細胞潛能」或「增加發育潛能」指增加細胞潛能或使細胞去分化為較少分化狀態的方法。例如,與非重編程狀態中的相同細胞相比較,具有增加的細胞潛能的細胞具有更多發育可塑性(即可分化成更多細胞類型)。換言之,重編程細胞是處於比非重編程狀態中的相同細胞較少分化狀態的細胞。
如本文使用的,術語「潛能」指細胞可接近的所有發育選項總和(即發育潛能)。本領域普通技術人員將認識到細胞潛能是連續體,範圍從最多可塑性細胞(具有最多發育潛能的全能幹細胞)到最少可塑性細胞(具有最少發育潛能的終末分化細胞)。細胞潛能的連續體包括但不限於全能細胞、多能細胞、多潛能細胞、寡能細胞、單能細胞和終末分化細胞。
如本文使用的,術語「多能的」指細胞形成機體或軀體(即胚體)的所有譜系的能力。例如,胚胎幹細胞是能夠形成來自三個胚層各自的細胞的一類多能幹細胞:外胚層、中胚層和內胚層。
多能性可部分通過評價細胞的多能性特徵進行測定。多能性特徵包括但不限於:(i)多能幹細胞形態;(ii)無限自我更新的潛能,(iii)多能幹細胞標記物包括但不限於下述的表達:SSEA1(僅小鼠)、SSEA3/4;SSEA5、TRA1-60/81;TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD90、CD105、OCT4、NANOG、SOX2、CD30和/或CD50;(iv)分化成所有三個體細胞譜系(外胚層、中胚層和內胚層)的能力,(v)由三個體細胞譜系組成的畸胎瘤形成;和(vi)由來自三個體細胞譜系的細胞組成的胚狀體形成。
兩類多能性先前已得到描述:類似於晚期胚泡的上胚層幹細胞(EpiSC)的多能性的「引發」或「亞穩」態,以及類似於早期/植入前胚泡的內細胞團的多能性的「幼稚」或「基」態。雖然兩種多能狀態均顯示出如上所述的特徵,但幼稚或基態還顯示出:(i)雌性細胞中的X染色體的預失活或復活,(ii)在單細胞培養期間改善的克隆形成能力和存活,(iii)DNA甲基化的總體減少,(iv)在發育調節基因啟動子上的H3K27me3阻遏染色質標記物沉積的減少,和(v)相對於引發態多能細胞,分化標記物的減少表達。其中外源多能性基因引入體細胞、表達且隨後沉默或從所得的多能細胞中去除的細胞重編程的標準方法一般可見具有多能性引發態的特徵。在標準多能細胞培養條件下,此類細胞保持引發態,除非外源轉基因表達得到維持,其中觀察到基態的特徵。
如本文使用的,術語「多能幹細胞形態」指胚胎幹細胞的典型形態特點。正常胚胎幹細胞形態的特徵在於圓且小的形狀、具有高核質比、核仁的顯著存在和典型細胞間間隔。
如本文使用的,術語「基因表達譜」、「基因表達標誌」、「基因表達板(panel)」、「基因板」或「基因標誌」指多重基因的表達或表達水平,所述基因作用於區分細胞或細胞群與另一細胞或細胞群。例如,在培養基中維持以防止自發分化的多能細胞群可展示基因表達譜,所述基因表達譜包含相對於並非在相同培養基中維持的相同起源的多能細胞對照群的分化基因的減少表達。
如本文使用的,術語「分化標記物基因」或「分化基因」指其表達指示在細胞例如多能細胞內發生的細胞分化的基因。分化標記物基因包括但不限於下述基因:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
如本文使用的,術語「分化標記物基因譜」、或「分化基因譜」、「分化基因表達譜」、「分化基因表達標誌」、「分化基因表達板」、「分化基因板」或「分化基因標誌」指多重分化標記物基因的表達或表達水平。
在特定實施例中,顯示減少的自發分化的多能細胞群的特徵可在於分化標記物基因或分化標記物基因譜的表達的減少。例如,可指示多能細胞或多能細胞群中減少的自發分化,其中與缺乏相同培養條件的對照多能細胞或多能細胞群的分化標記物基因表達相比較,給定組的培養條件引起一種或多種分化標記物基因表達的至少10%、20%、25%、30%、40%、50%、60%、70%、80%、90%或更多的減少。
如本文使用的,「基因表達」指生物樣品例如多能細胞或包含多能細胞的細胞群中的基因的表達和/或表達模式的相對水平。在特定實施例中,多能細胞是iPSC。
本文涵蓋本領域可得的用於檢測表徵本發明細胞的基因表達的任何方法。如本文使用的,術語「檢測表達」意指測定基因的RNA轉錄物或其表達產物的數量或存在。用於檢測基因表達的方法,即基因表達概況分析,包括基於多核苷酸的雜交分析的方法、基於多核苷酸測序的方法、免疫組織化學方法和基於蛋白質組學的方法。方法一般檢測目的基因的表達產物(例如mRNA)。在一些實施例中,使用基於PCR的方法例如逆轉錄PCR(RT-PCR)(Weis等人,TIG 8:263-64,1992),以及基於陣列的方法例如微陣列(Schena等人,Science 270:467-70,1995)。
「貼壁」指在適當培養基的存在下,細胞附著至容器,例如細胞附著至無菌塑料(或塗布塑料)細胞培養皿或燒瓶。某些類別的細胞在培養中無法持續或不生長,除非它們貼壁至細胞培養容器。某些類別的細胞(「非貼壁細胞」)無需貼壁在培養中維持和/或增殖。
「培養」或「細胞培養」指在體外環境中的細胞維持、生長和/或分化。「細胞培養基」、「培養基」(在每種情況下單數的「培養基」)、「補充物」和「培養基補充物」指栽培細胞培養物的營養組合物。
「栽培」指在組織或機體外,例如在無菌塑料(或塗布塑料)細胞培養皿或燒瓶中的細胞持續、繁殖(生長)和/或分化。「栽培」可利用培養基作為營養素、激素和/或幫助使細胞繁殖和/或持續的其他因子的源。
如本文使用的,「解離的」細胞指已與其他細胞或表面(例如培養板表面)基本上分開或者純化離開其他細胞的細胞。例如,細胞可通過機械或酶促方法與動物或組織解離。可替代地,在體外聚集的細胞可酶促或機械地例如通過解離成簇、單細胞或單細胞和簇的混合物的懸浮液彼此解離。在另外一個可替代實施例中,貼壁細胞與培養板或其他表面解離。解離因此可涉及破壞細胞與細胞外基質(ECM)和基質(例如培養表面)的相互作用,或破壞細胞之間的ECM。
如本文使用的,術語「富集(enrich)」和「富集(enriching)」指增加組合物例如細胞組合物中的指定組分的量,並且當用於描述細胞的組合物例如細胞群時,「富集的」指與富集前細胞群中的此類組分的比例相比較,具有按比例增加量的指定組分的細胞群。例如,組合物例如細胞群可就靶細胞類型(即具有指定特徵的細胞)而言進行富集,因此與富集前細胞群中存在的靶細胞比例相比較,具有增加比例或百分比的靶細胞類型。細胞群可通過本領域已知的細胞選擇和分選方法就靶細胞類型進行富集。在一些實施例中,細胞群通過如本文實例中所述的分選或選擇方法進行富集。在一個特定實施例中,就靶細胞群富集的方法使細胞群就靶細胞群而言富集至少約20%,從而意指富集的細胞群包含比群富集前群中按比例多約20%的靶細胞類型。在一個實施例中,就靶細胞群富集的方法使細胞群就靶細胞群而言按比例富集至少約30+%、40+%、50+%、60+%、70+%、80%、85%、90%、95%、97%、98%或99%,或至少約98%,或在特定實施例中,約99%。
在某些實施例中,細胞群就多能細胞或顯示出多能性特徵的細胞的量而言進行富集。在本發明的特定實施例中,經歷重編程的細胞群就具有多能性特徵的靶細胞進行富集,所述多能性特徵例如表達多能性標記物包括但不限於SSEA3、SSEA4、TRA 1-60、TRA-1-81、CD30或CD50。
在特定實施例中,使用對於分化細胞譜系或非多能細胞特異性的表面標記物,使細胞群例如經歷重編程的細胞群耗盡非多能細胞,所述表面標記物可包括例如CD13、CD26、CD34、CD45、CD31、CD46或CD7。所得到的細胞群因此可描述為富含多能細胞的細胞群。
在特定實施例中,富集細胞包含不同的基因或蛋白質表達譜,例如至少兩種多能性標記物例如SSEA3、SSEA4、TRA 1-60、TRA-1-81、CD30和CD50的細胞表面表達。在一些實施例中,富集細胞包含兩種或更多種多能性標記物。在特定實施例中,富集細胞表達與TRA-181或TRA-160組合的SSEA4。在更特定的實施例中,富集細胞表達SSEA4、TRA181和CD30。在一個實施例中,細胞群包含至少約5%、10%、15%、20%、25%、30%、40%、50%、70%、75%、80%、90%、95%、97%、98%或99%的富集細胞例如多能細胞。
因此,在一些實施例中,就多能細胞富集細胞群的方法包括基於多能性標記物例如SSEA3、SSEA4、TRA 1-60、TRA-1-81、CD30和CD50的細胞表面表達分選細胞群,並且收集表達此類標記物的細胞級分,以獲得富含多能細胞的細胞群。在其他實施例中,通過基於分化中或分化細胞標記物例如CD13、CD26、CD34、CD45、CD31、CD46和CD7的細胞表面表達分選細胞群,並且耗盡此類細胞的細胞群以獲得富含多能細胞的細胞群,使細胞群富含多能細胞。在特定實施例中,細胞群基於CD13的表達進行分選,並且從細胞群中去除CD13+細胞,以獲得富含多能細胞的細胞群。
如本文使用的,「飼養細胞」或「飼養者」是用於描述與第二類型的細胞共培養的一個類型的細胞,以提供第二類型的細胞可在其中生長的環境,因為飼養細胞提供用於支持第二細胞類型的生長因子和營養素。飼養細胞任選來自與它們支持的細胞不同的物種。例如,某些類型的人細胞包括幹細胞可通過小鼠胚胎成纖維細胞和永生化小鼠胚胎成纖維細胞的原代培養物得到支持。當與其他細胞共培養時,飼養細胞通常可通過照射或用抗有絲分裂劑例如絲裂黴素c處理而失活,以防止其生長超過它們支持的細胞。不限於前文,一個具體飼養細胞類型可為人飼養者,例如人皮膚成纖維細胞。另一飼養細胞類型可以是小鼠胚胎成纖維細胞(MEF)。
如本文使用的,「無飼養者」(FF)環境指大體上不含飼養細胞和/或未通過飼養細胞栽培而預條件化的環境例如細胞培養或培養基。「預條件化的」培養基指在飼養細胞已在培養基內栽培一段時間例如至少一天後收穫的培養基。預條件化的培養基含有許多介質物質,包括通過在培養基中栽培的飼養細胞分泌的生長因子和細胞因子。
基因組穩定性指細胞忠實複製DNA且維持DNA複製過程的完整性的能力。如本文使用的,術語「基因組穩定的細胞」和「具有基因組穩定性的細胞」指顯示出一定頻率的突變和染色體異常(例如易位、非整倍體、拷貝數變異和重複)的細胞,所述頻率基本上類似於相對於正常人體細胞的突變和染色體異常的頻率。
「成分」指可用於細胞培養基中以維持和/或促進細胞生長和/或分化的任何化合物或其他材料,無論在起源中是化學還是生物的。術語「組分」、「營養素」和「成分」可互換使用。用於細胞培養基的常規成分可包括但不限於胺基酸、鹽、金屬、糖、脂質、核酸、激素、維生素、脂肪酸、蛋白質等等。促進和/或維持離體細胞栽培的其他成分可通過本領域普通技術人員根據需要對於所需效應加以選擇。
「分離(isolate)」或「分離(isolating)」指使組合物或材料與其天然環境分開且收集,例如個別細胞或細胞培養物與組織或機體的分開。在一個方面,細胞群或組合物基本上不含它在自然界中可與之結合的細胞和材料。就靶細胞群而言,「分離的」或「純化的」或「基本上純的」指就構成總細胞群的靶細胞而言,至少約50%、至少約75%、至少約85%、至少約90%,並且在特定實施例中,至少約95%純的細胞群。細胞群或組合物的純度可通過本領域眾所周知的適當方法進行評價。例如,基本上純的多能細胞群指就構成總細胞群的多能細胞而言,至少約50%、至少約75%、至少約85%、至少約90%,並且在特定實施例中,至少約95%,並且在某些實施例中,約98%純的細胞群。術語「大體上純的」在本文中可與「基本上純的」互換使用。
「傳代(passage)」或「傳代(passaging)」指當細胞已增殖至所需程度時,將細胞細分且鋪平板到多重細胞培養表面或容器內的動作。在一些實施例中,「傳代(passage)」或「傳代(passaging)」指將細胞細分、稀釋且鋪平板。當細胞從原代培養表面或容器傳代到後續組的表面或容器內時,後續培養在本文中可被稱為「繼代培養」或「第一次傳代」等。每個細分且鋪平板到新培養容器內的動作被視為一次傳代。
「鋪平板」指將一個或多個細胞置於培養容器內,使得細胞貼壁至細胞培養容器且在細胞培養容器上鋪展。
「多能性因子」指單獨或與其他試劑組合能夠增加細胞的發育潛能的試劑。多能性因子包括但不限於能夠增加細胞的發育潛能的多核苷酸、多肽和小分子。示例性多能性因子包括例如轉錄因子和小分子重編程試劑。
「增殖」指一個細胞分裂成兩個大體上等同的細胞或細胞群數目增加(例如複製)的性質。
「繁殖」指使組織或機體外的細胞例如在無菌容器例如塑料(或塗布塑料)細胞培養皿或燒瓶中生長(例如經由細胞增殖複製)。
「原代培養」指其中經分離的細胞置於具有培養基的第一培養容器中的細胞、組織和/或培養物。然而,細胞、組織和/或培養物可持續和/或可增殖,只要細胞、組織和/或培養物保留在第一容器中,細胞、組織和/或培養物被稱為原代培養。
術語「小分子重編程試劑」或「小分子重編程化合物」在本文中可互換使用,並且指單獨或與其他多能性因子組合可增加細胞的發育潛能的小分子。「小分子」指具有小於約5kD、小於約4kD、小於約3kD、小於約2kD、小於約1kD或小於約.5kD的分子量的試劑。小分子包括但不限於:核酸、擬肽、類肽、碳水化合物、脂質或者其他有機或無機分子。化學和/或生物混合物例如真菌、細菌或藻類提取物的文庫是本領域已知的,並且在某些實施例中,可用作小分子源。在特定實施例中,本文使用的小分子重編程試劑具有小於10,000道爾頓,例如小於8000、6000、4000、2000道爾頓,例如在50-1500、500-1500、200-2000、500-5000道爾頓之間的分子量。
C.細胞
在一個特定實施例中,一個或多個細胞可使用本文考慮的組合物和方法進行培養、解離且傳代。在一個實施例中,單細胞使用本文考慮的組合物和方法進行培養、解離且傳代。在另一個實施例中,細胞群或多個細胞使用本文考慮的組合物和方法進行培養、解離且傳代。
適用於特定實施例中的起始細胞群可來源於大體上任何合適的源,並且就細胞類型或多能性狀態而言可以是異質或同質的。合適的細胞包括胎兒細胞和成人細胞。另外,合適的細胞在起源中可以是哺乳動物,例如來自嚙齒類動物、貓、犬、豬、山羊、綿羊、馬、牛或靈長類動物。在一個實施例中,細胞是人細胞。
細胞可以是體細胞、非多能、不完全或部分多能幹細胞、多潛能細胞、寡能細胞、單能細胞、終末分化細胞、或包含前述的任何組合的混合細胞群。適用於特定實施例中的多能細胞包括但不限於天然存在的幹細胞、胚胎幹細胞或iPSC。「混合」細胞群是具有不同程度的發育潛能的細胞群。例如,混合細胞群可包含經歷重編程的細胞,使得混合群包含多能細胞、部分多能細胞和非多能細胞例如完全分化細胞。
在一個實施例中,起始細胞群選自成人或新生兒幹/祖細胞。在特定實施例中,起始幹/祖細胞群選自:中胚層幹/祖細胞、內胚層幹/祖細胞和外胚層幹/祖細胞。
中胚層幹/祖細胞的舉例說明性例子包括但不限於:中胚層幹/祖細胞、內皮幹/祖細胞、骨髓幹/祖細胞、臍帶幹/祖細胞、脂肪組織衍生的幹/祖細胞、造血幹/祖細胞(HSC)、間充質幹/祖細胞、肌肉乾/祖細胞、腎幹/祖細胞、成骨細胞幹/祖細胞、軟骨細胞幹/祖細胞等等。
外胚層幹/祖細胞的舉例說明性例子包括但不限於神經幹/祖細胞、視網膜幹/祖細胞、皮膚幹/祖細胞等等。
內胚層幹/祖細胞的舉例說明性例子包括但不限於肝幹/祖細胞、胰腺幹/祖細胞、上皮幹/祖細胞等等。
在某些實施例中,起始細胞群可以是選自下述的異質或同質細胞群:胰島細胞、CNS細胞、PNS細胞、心肌細胞、骨骼肌細胞、平滑肌細胞、造血細胞、骨細胞、肝細胞、脂肪細胞、腎細胞、肺細胞、軟骨細胞、皮膚細胞、濾泡細胞、血管細胞、上皮細胞、免疫細胞、內皮細胞等等。
D.用於減少自發分化和誘導基態多能性的培養平臺
細胞建庫、疾病建模和細胞療法應用已對製造高質量多能細胞提出漸增需求。例如,無足跡iPSC的高流通量衍生及其在允許規模化生產的系統中的擴增仍是技術上難以捉摸的。在特定實施例中,考慮了允許在階段特異性培養基組合物中使用小分子途徑抑制劑的多能細胞的快速、平行生成、選擇和擴增的培養平臺。本文考慮的平臺支持在完全無飼養者環境中使用最低限度重編程因子的有效和加快的重編程;使多能細胞的單細胞培養和擴增成為可能,同時維持同質和基因組穩定的多能群。此外,本文考慮的培養平臺提供了將多能細胞包括hESC和hiPSC培養至減少的自發分化狀態和常見的基態多能性,與遺傳背景無關且不依賴於轉基因表達。
本文考慮的培養平臺可部分用於產生在培養中具有減少的自發分化的工業或臨床級別的多能細胞。在一個實施例中,非多能細胞進行誘導,以變成多能細胞,並且培養以維持多能性。在另一個實施例中,非多能細胞進行誘導,以變成多能細胞,並且培養以實現和/或維持培養中減少的自發分化。在另一個實施例中,非多能細胞進行誘導,以變成多能細胞,並且培養以實現和/或維持基態多能性。
在各個實施例中,本文考慮的培養平臺使一個或多個多能細胞維持基態多能性、正常核型和基因組穩定性至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、60、70、80、90、100次或更多次傳代,包括任何中間數目的傳代。
在其他實施例中,本文考慮的培養平臺使一個或多個多能性細胞中減少的自發分化維持至少1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、60、70、80、90、100次或更多次傳代,包括任何中間數目的傳代。
在一個實施例中,培養平臺包括細胞培養基,所述細胞培養基包含細胞培養基和GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑。在各個實施例中,本文考慮的細胞培養基不包含或缺乏TGFβ/激活素信號傳導途徑的抑制劑,包括TGFβ受體(TGFβR)抑制劑和ALK5抑制劑。不希望受任何特定理論束縛,本發明人驚訝地發現雖然TGFβR/ALK5抑制劑增加重編程效率,但這些抑制劑抵消多能細胞群的長期維持、質量和同質性,即TGFβ途徑信號傳導的抑制改善細胞重編程的效率,但從這種抑制中解除是多能細胞群在體外培養系統中,特別是在使用無飼養細胞和單細胞、酶促傳代的系統中的後續維持所需的,其中具有減少的自發分化的同質多能群是優選的,並且更特別地,其中不存在轉基因表達。另外,如本文公開的,在包含GSK-3抑制劑和MEK抑制劑和任選的ROCK抑制劑,但缺乏TGFβR/ALK5抑制劑的培養基中培養亞穩多能細胞,使多能細胞轉變,以實現減少的自發分化和/或實現基態多能性。本文考慮的培養基平臺還使多能細胞在無飼養者環境中的有效重編程和長期培養成為可能。此外,雖然「ALK5抑制劑」不預期涵蓋非特異性激酶抑制劑,但「ALK5抑制劑」應理解為涵蓋除ALK5之外還抑制ALK4和/或ALK7的抑制劑,例如SB-431542(參見例如Inman等人,J Mol.Pharmacol.62(1):65-74(2002))。
在一個優選實施例中,培養平臺包括包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和任選的LIF和/或bFGF的細胞培養基,並且不包含TGFβ/激活素信號傳導途徑的小分子抑制劑,包括但不限於TGFβR或ALK5抑制劑。
在另外的實施例中,細胞培養基基本上不含細胞因子和/或生長因子,且任選是無飼養者環境。在其他實施例中,細胞培養基含有補充物例如血清、提取物、生長因子、激素、細胞因子等等。
在一個優選實施例中,培養平臺包含無飼養者培養。
本文考慮的培養平臺還提供眾多優點,例如製造具有減少的自發分化和/或實現基態多能性的工業或臨床級別多能細胞的同質群。如本文使用的,術語「同質的」指其中每個細胞與群中的其他細胞相同或基本上相同的細胞群。在一個實施例中,如果每個細胞表達與本文考慮的相同多能性標記物(例如SSEA4和TRA1-81)中的一種或多種,則細胞與群中的其他細胞相同。在一個實施例中,如果至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或更多細胞與群中的其他細胞相同或基本上相同,則群是同質的。
1.TGFβ受體/ALK5抑制劑
TGFβ受體(例如ALK5)抑制劑可包括針對TGFβ受體(例如ALK5)的抗體、TGFβ受體的顯性失活(negative)變體和壓制TGFβ受體表達的反義核酸。示例性TGFβ受體/ALK5抑制劑包括但不限於SB431542(參見例如Inman等人,Molecular Pharmacology 62(1):65-74(2002)),A-83-01也稱為3-(6-甲基-2-吡啶基)-N-苯基-4-(4-喹啉基)-1H-吡唑-1-硫代甲醯胺(參見例如Tojo等人,Cancer Science 96(11):791-800(2005),且由例如Toicris Bioscience商購可得);2-(3-(6-甲基吡啶-2-基)-1H-吡唑-4-基)-1,5-萘啶,Wnt3a/BIO(參見例如以引用的方式併入本文的Dalton等人,WO2008/094597),BMP4(參見Dalton,同上),GW788388(-{4-[3-(吡啶-2-基)-1H-吡唑-4-基]吡啶-2-基}-N-(四氫-2H-吡喃-4-基)苯甲醯胺)(參見例如Gellibert等人,Journal of Medicinal Chemistry 49(7):2210-2221(2006)),SM16(參見例如Suzuki等人,Cancer Research 67(5):2351-2359(2007)),IN-1130(3-((5-(6-甲基吡啶-2-基)-4-(喹喔啉-6-基)-1H-咪唑-2-基)甲基)苯甲醯胺)(參見例如Kim等人,Xenobiotica 38(3):325-339(2008)),GW6604(2-苯基-4-(3-吡啶-2-基-1H-吡唑-4-基)吡啶)(參見例如de Gouville等人,Drug News Perspective 19(2):85-90(2006)),SB-505124(2-(5-苯並[1,3]二氧雜環戊烯-5-基-2-叔丁基-3H-咪唑-4-基)-6-甲基吡啶鹽酸鹽)(參見例如DaCosta等人,Molecular Pharmacology 65(3):744-752(2004))和嘧啶衍生物(參見例如以引用的方式併入本文的Stiefl等人,WO2008/006583中列出的那些)。此外,雖然「ALK5抑制劑」不預期涵蓋非特異性激酶抑制劑,但「ALK5抑制劑」應理解為涵蓋除ALK5之外還抑制ALK4和/或ALK7的抑制劑,例如SB-431542(參見例如Inman等人,J Mol.Pharmacol.62(1):65-74(2002))。不預期限制本發明的範圍,認為ALK5抑制劑影響間充質至上皮轉換/轉變(MET)過程。TGFβ/激活素途徑是上皮至間充質轉變(EMT)的驅動物。因此,抑制TGFβ/激活素途徑可有助於MET(即重編程)過程。
考慮到本文顯示抑制ALK5的效應的數據,認為TGFβ/激活素途徑的抑制將具有抑制ALK5的相似效應。因此,TGFβ/激活素途徑的任何抑制劑(例如上遊或下遊)可與ALK5抑制劑組合使用或代替ALK5抑制劑使用,如本文的每個段落中所述。示例性TGFβ/激活素途徑抑制劑包括但不限於:TGFβ受體抑制劑、SMAD 2/3磷酸化抑制劑、SMAD 2/3和SMAD 4相互作用抑制劑、以及SMAD 6和SMAD 7激活物/激動劑。此外,下文描述的分類僅用於組構目的,並且本領域技術人員將知道化合物可影響途徑內的一個或多個點,並且因此化合物可在超過一個限定類別中起作用。
TGFβ受體(TGFβR)抑制劑可包括針對TGFβ受體的抗體、TGFβ受體的顯性失活變體以及靶向TGFβ受體的siRNA或反義核酸。TGFβ受體抑制劑的具體例子包括但不限於SU5416;2-(5-苯並[1,3]二氧雜環戊烯-5-基-2-叔丁基-3H-咪唑-4-基)-6-甲基吡啶鹽酸鹽(SB-505124);lerdelimumb(CAT-152);美替木單抗(CAT-192);GC-1008;ID11;AP-12009;AP-11014;LY550410;LY580276;LY364947;LY2109761;SB-505124;SB-431542;SD-208;SM16;NPC-30345;Ki26894;SB-203580;SD-093;Gleevec;3,5,7,2',4'-五羥基黃酮(Morin);激活素-M108A;P144;可溶性TBR2-Fc;以及靶向TGFβ受體的反義轉染的腫瘤細胞。(參見例如Wrzesinski等人,Clinical Cancer Research 13(18):5262-5270(2007);Kaminska等人,Acta Biochimica Polonica 52(2):329-337(2005);和Chang等人,Frontiers in Bioscience 12:4393-4401(2007))。
SMAD 2/3磷酸化抑制劑可包括針對SMAD2或SMAD3的抗體、SMAD2或SMAD3的顯性失活變體、以及靶向SMAD2或SMAD3的反義核酸。抑制劑的具體例子包括PD169316;SB203580;SB-431542;LY364947;A77-01;和3,5,7,2',4'-五羥基黃酮(Morin)。(參見例如以引用的方式併入本文的Wrzesinski,同上;Kaminska,同上;Shimanuki等人,Oncogene 26:3311-3320(2007);和Kataoka等人,EP1992360)。
SMAD 2/3和smad4相互作用抑制劑可包括針對SMAD2、SMAD3和/或smad4的抗體,SMAD2、SMAD3和/或smad4的顯性失活變體,以及靶向SMAD2、SMAD3和/或smad4的反義核酸。SMAD 2/3和smad4相互作用抑制劑的具體例子包括但不限於Trx-SARA、Trx-xFoxH1b和Trx-Lef1。(參見例如Cui等人,Oncogene 24:3864-3874(2005)和Zhao等人,Molecular Biology of the Cell,17:3819-3831(2006))。
SMAD 6和SMAD 7激活物/激動劑包括但不限於針對SMAD 6或SMAD 7的抗體,SMAD 6或SMAD 7的顯性失活變體,以及靶向SMAD 6或SMAD 7的反義核酸。抑制劑的具體例子包括但不限於smad7-as PTO-寡核苷酸。(參見例如兩者均以引用的方式併入本文的Miyazono等人,US6534476,以及Steinbrecher等人,US2005119203)。
2.WNT途徑激動劑
如本文使用的,術語「Wnt信號促進試劑」、「Wnt途徑激活試劑」或「Wnt途徑激動劑」指Wnt信號傳導途徑的激動劑,包括但不限於Wntl、Wnt2、Wnt2b/13、Wnt3、Wnt3a、Wnt4、Wnt5a、Wnt5b、Wnt6、Wnt7a、Wnt7b、Wnt7c、Wnt8、Wnt8a、Wnt8b、Wnt8c、Wnt10a、Wntl0b、Wnt11、Wnt14、Wnt15或Wnt16中的一種或多種的激動劑。Wnt途徑激動劑還包括但不限於下述多肽或其片段中的一種或多種:Dkk多肽、crescent多肽、cerberus多肽、axin多肽、Frzb多肽、T細胞因子多肽、或顯性失活散亂(disheveled)多肽。
Wnt途徑激動劑的非限制性例子還包括下述中的一種或多種:包含編碼Wnt多肽的核苷酸序列的核酸,包含Wnt多肽的胺基酸序列的多肽,包含編碼活化Wnt受體的核苷酸序列的核酸,包含活化Wnt受體的胺基酸序列的多肽,促進Wnt/β-連環蛋白信號傳導的小有機分子,抑制Wnt拮抗劑的表達或活性的小有機分子,抑制Wnt拮抗劑表達的反義寡核苷酸,抑制Wnt拮抗劑表達的核酶,抑制Wnt拮抗劑表達的RNAi構建體、siRNA或shRNA,與Wnt拮抗劑結合且抑制Wnt拮抗劑活性的抗體,包含編碼β-連環蛋白多肽的核苷酸序列的核酸,包含β-連環蛋白多肽的胺基酸序列的多肽,包含編碼Lef-1多肽的核苷酸序列的核酸,包含Lef-1多肽的胺基酸序列的多肽。
Wnt途徑激動劑還包括GSK3抑制劑,例如包含編碼顯性失活GSK-3、GSK3α或GSK3β多肽的核苷酸序列的核酸,包含顯性失活GSK-3、GSK3α或GSK3β多肽的胺基酸序列的多肽,與GSK-3、GSK3α或GSK3β結合且抑制GSK-3、GSK3α或GSK3β的表達或活性的小有機分子,與GSK-3、GSK3α或GSK3β結合且抑制GSK-3、GSK3α或GSK3β的表達和/或活性的RNAi構建體、siRNA或shRNA,與GSK-3、GSK3α或GSK3β結合且抑制GSK-3、GSK3α或GSK3β的表達的反義寡核苷酸,與GSK-3、GSK3α或GSK3β結合且抑制GSK-3、GSK3α或GSK3β的表達和/或活性的抗體,與GSK-3、GSK3α或GSK3β結合且抑制GSK-3、GSK3α或GSK3β的表達的核酶,以及在效應上類似於GSK3抑制的活化β-連環蛋白靶基因的任何GSK-3不依賴性試劑。
3.GSK-3β抑制劑
GSK-3β抑制劑是適用於本文考慮的組合物中的具體示例性Wnt途徑激動劑,並且可包括但不限於多核苷酸、多肽和小分子。本文考慮的GSK-3β抑制劑可減少GSK-3β表達和/或GSK-3β活性。本文考慮的GSK-3β抑制劑的舉例說明性例子包括但不限於抗GSK-3β抗體,顯性失活GSK-3β變體,靶向GSK-3β的siRNA、shRNA、miRNA和反義核酸。
其他舉例說明性GSK-3β抑制劑包括但不限於:肯帕羅酮、1-氮雜坎帕羅酮、CHIR99021、CHIR98014、AR-A014418、CT 99021、CT 20026、SB216763、AR-A014418、鋰、SB 415286、TDZD-8、BIO、BIO-丙酮肟、(5-甲基-1H-吡唑-3-基)-(2-苯基喹唑啉-4-基)胺、吡啶卡巴唑-環戊二烯基釕絡合物、TDZD-8 4-苄基-2-甲基-1,2,4-噻二唑烷-3-,5-二酮、2-硫代(3-碘苄基)-5-(1-吡啶基)-[1,3,4]-噁二唑、OTDZT、α-4-二溴苯乙酮、AR-AO 144-18、3-(1-(3-羥丙基)-1H-吡咯並[2,3-b]吡啶-3-基]-4-吡嗪-2-基-吡咯-2,5-二酮;TWSl 19吡咯並嘧啶化合物、L803H-KEAPPAPPQSpP-NH2或其肉豆蔻醯化形式;2-氯-1-(4,5-二溴-噻吩-2-基)-乙酮;GF109203X;RO318220;TDZD-8;TIBPO;和OTDZT。
在特定舉例說明性實施例中,GSK-3β抑制劑是CHIR99021、BIO或肯帕羅酮。
在一個優選實施例中,GSK-3β抑制劑是CHIR99021。
4.ERK/MEK抑制劑
適用於本文考慮的組合物中的ERK/MEK抑制劑包括但不限於多核苷酸、多肽和小分子。本文考慮的ERK/MEK抑制劑可減少MEK或ERK表達和/或MEK或ERK活性。本文考慮的ERK/MEK抑制劑的舉例說明性例子包括但不限於抗MEK或抗ERK抗體,顯性失活MEK或ERK變體,靶向MEK或ERK的siRNA、shRNA、miRNA和反義核酸。
其他舉例說明性ERK/MEK抑制劑包括但不限於PD0325901、PD98059、UO126、SL327、ARRY-162、PD184161、PD184352、舒尼替尼、索拉非尼、凡德他尼、帕唑帕尼、阿西替尼、GSKl 120212、ARRY-438162、RO5126766、XL518、AZD8330、RDEAl 19、AZD6244、FR180204和PTK787。
另外的舉例說明性MEK/ERK抑制劑包括公開於國際公開專利申請WO 99/01426、WO 02/06213、WO 03/077914、WO 05/051301和WO2007/044084中的那些化合物。
MEK/ERK抑制劑的進一步舉例說明性例子包括下述化合物:6-(4-溴-2-氯-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-羧酸(2,3-二羥基-丙氧基)-醯胺;6-(4-溴-2-氯-苯基氨基)-7-氟-3-(四氫-吡喃-2-基甲基)-3H-苯並咪唑-5-羧酸(2-羥基-乙氧基)-醯胺、1-[6-(4-溴-2-氯-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-基]-2-羥基乙酮、6-(4-溴-2-氯-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-羧酸(2-羥基-1,1-二甲基-乙氧基)-醯胺、6-(4-溴-2-氯-苯基氨基)-7-氟-3-(四氫-呋喃-2-基甲基)-3H-苯並咪唑-5-羧酸(2-羥基-乙氧基)-醯胺、6-(4-溴-2-氟-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-羧酸(2-羥基-乙氧基)-醯胺、6-(2,4-二氯-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-羧酸(2-羥基-乙氧基)-醯胺、6-(4-溴-2-氯-苯基氨基)-7-氟-3-甲基-3H-苯並咪唑-5-羧酸(2-羥基-乙氧基)-醯胺,下文被稱為MEK抑制劑1;2-[(2-氟-4-碘苯基)氨基]-N-(2-羥基乙氧基)-1,5-二甲基-6-氧代-1,6-二氫吡啶-3-甲醯胺;下文被稱為MEK抑制劑2;和4-(4-溴-2-氟苯基氨基)-N-(2-羥基乙氧基)-1,5-二甲基-6-氧代-1,6-二氫噠嗪-3-甲醯胺或其藥學可接受的鹽。
在一個優選實施例中,MEK/ERK抑制劑是PD98059。
5.ROCK抑制劑
Rho相關激酶(ROCK)是充當Rho激酶的下遊效應物的絲氨酸/蘇氨酸激酶(其中存在三種同種型--RhoA、RhoB和RhoC)。適用於本文考慮的組合物中的ROCK抑制劑包括但不限於多核苷酸、多肽和小分子。本文考慮的ROCK抑制劑可減少ROCK表達和/或ROCK活性。本文考慮的ROCK抑制劑的舉例說明性例子包括但不限於抗ROCK抗體,顯性失活ROCK變體,靶向ROCK的siRNA、shRNA、miRNA和反義核酸。
本文考慮的舉例說明性ROCK抑制劑包括但不限於:thiazovivin、Y27632、Fasudil、AR122-86、Y27632H-1152、Y-30141、Wf-536、HA-1077、羥基-HA-1077、GSK269962A、SB-772077-B、N-(4-吡啶基)-N'-(2,4,6-三氯苯基)脲、3-(4-吡啶基)-1H-吲哚、和(R)-(+)-反式-N-(4-吡啶基)-4-(1-氨基乙基)-環己烷甲醯胺以及公開於美國專利號8,044,201中的ROCK抑制劑,所述美國專利以引用的方式全文併入本文。
在一個實施例中,ROCK抑制劑是thiazovivin、Y27632或pyrintegrin。
在一個優選實施例中,ROCK抑制劑是thiazovivin。
在本文考慮的組合物和細胞培養基中的小分子的量可改變,並且可根據具體培養條件進行優化,包括所使用的具體分子和組合、在培養基中培養的細胞類型以及具體應用。在一個實施例中,小分子存在於組合物中的濃度足以誘導多能性、改善重編程效率、增加或維持細胞的潛能、或者誘導或維持基態多能性。
在特定實施例中,在本發明的細胞培養基中的小分子的優選濃度和組合在表1中顯示為命運維持培養基(FMM)。培養基的組分可以在表1中所示的最佳濃度附近的最佳範圍內的量存在於培養基中。命運重編程培養基(FRM)可用於包括細胞重編程的本文考慮的培養平臺中,但不適合於基態多能細胞的建立和長期維持。
表1
6.細胞因子和生長因子
在特定實施例中,本發明的細胞培養基基本上不含細胞因子和/或生長因子。在某些實施例中,細胞培養基含有一種或多種補充物,包括但不限於血清、提取物、生長因子、激素、細胞因子等等。
在一個舉例說明性實施例中,培養基可包含ECM蛋白、層粘連蛋白1、纖連蛋白、IV型膠原同種型、蛋白酶、蛋白酶抑制劑、細胞表面粘附蛋白、細胞信號傳導蛋白質、鈣粘素、氯離子細胞內通道1、跨膜受體PTK7、胰島素樣生長因子、或抑制素βA中的一種或多種,但不包含TGFβ/激動素/nodal信號傳導途徑的誘導物和激活素A。在其他實施例中,培養基可包含TGFβ/激動素/nodal信號傳導途徑的誘導物。
在另一個舉例說明性實施例中,培養基包含下述細胞因子或生長因子中的一種或多種:表皮生長因子(EGF)、酸性成纖維細胞生長因子(aFGF)、鹼性成纖維細胞生長因子(bFGF)、白血病抑制因子(LIF)、肝細胞生長因子(HGF)、胰島素樣生長因子-1(IGF-1)、胰島素樣生長因子2(IGF-2)、角化細胞生長因子(KGF)、神經生長因子(NGF)、血小板衍生的生長因子(PDGF)、轉化生長因子β(TGF-β)、血管內皮細胞生長因子(VEGF)、轉鐵蛋白、各種白介素(如IL-1至IL-18)、各種集落刺激因子(如粒細胞/巨噬細胞集落刺激因子(GM-CSF))、各種幹擾素(如IFN-γ)以及對幹細胞具有作用的其他細胞因子例如幹細胞因子(SCF)和促紅細胞生成素(Epo)。這些細胞因子可在商業上得自例如R&D Systems,Minneapolis,Minn,並且可為天然的或重組的。在特定實施例中,生長因子和細胞因子可以本文考慮的濃度添加。在某些實施例中,生長因子和細胞因子可以憑經驗測定或如由確認的細胞因子技術指導的濃度添加。
7.培養基質
任何合適的容器或細胞培養容器均可用作基礎培養基和/或細胞培養補充物中的細胞培養的支持物。在支持物上的基質塗層不是必要的。然而,用貼壁促進基質(例如膠原、纖連蛋白、含RGD多肽、明膠等等)塗布培養容器的表面促進細胞的附著,並且在特定實施例中,可增強本文公開的細胞培養基和補充物的效應。用於培養和傳代細胞的合適基質是本領域已知的,並且包括但不限於玻連蛋白、明膠、層粘連蛋白、纖連蛋白、膠原、彈性蛋白、骨橋蛋白、天然存在的細胞系生產的基質的混合物例如MatrigelTM以及合成或人造表面例如聚胺單層和羧基封端單層。
在一個實施例中,本文考慮的培養平臺包括包含MatrigelTM或玻連蛋白的基質。
8.無飼養者環境
用於培養多能細胞的現有方法在很大程度上依賴於飼養細胞或用飼養細胞預條件化且含有胎牛血清的培養基;然而,此類環境可能不適合於產生用於臨床和治療用途的細胞。例如,在此類異物汙染的環境中栽培的細胞一般視為不適合於人細胞移植,因為對動物組分的暴露可呈現免疫排斥和將未經鑑定的病原體傳遞給被治療患者的嚴重危險,並且可潛在使動物逆轉錄病毒復活。使用無動物培養基例如本文考慮的無飼養者環境的培養系統有助於臨床級別細胞系特別是hESC和hiPSC細胞系的製造。
在特定實施例中,無飼養者環境基本上不含人飼養細胞,包括但不限於小鼠胚胎成纖維細胞、人成纖維細胞、角化細胞和胚胎幹細胞,並且不通過飼養細胞預條件化。無飼養者細胞培養基適用於培養多能細胞,使細胞重編程,多能細胞的單細胞培養、解離和傳代,多能細胞的細胞分選,基態多能細胞的生成,以及基態多能性的維持。在特定實施例中,無飼養者環境用於誘導多能性,改善重編程效率,和/或增加或維持細胞潛能。在某些實施例中,無飼養者環境基本上不含細胞因子和生長因子,包括bFGF。
9.解離
由本文考慮的培養平臺提供的優點之一在於培養、傳代和解離單個基態多能細胞的增強活力和存活。細胞解離成單細胞例如單細胞懸浮液可通過酶促或機械手段來完成。本領域已知的允許細胞解離成單細胞的任何酶促試劑均可用於本發明的方法中。在一個實施例中,解離試劑選自胰蛋白酶/EDTA、TrypLE-Select、膠原酶IV和分散酶。
根據本文考慮的方法,在解離細胞中還可使用單獨或與酶促試劑組合的螯合劑例如EDTA、Accutase或AccuMax。解離試劑可溶解於無鈣和鎂的PBS中,以有助於解離為單細胞。
為了增強細胞在解離期間和在解離後的存活,可加入存活促進物質(例如生長因子、涉及細胞死亡和細胞凋亡的細胞途徑的抑制劑、或條件化培養基),例如ROCK抑制劑例如thiazovivin。
細胞培養和培養基收集中的技術在下述中概述:Hu等人,Curr.Opin.Biotechnol.8:148,1997;K.Kitano,Biotechnology 17:73,1991;Curr.Opin.Biotechnol.2:375,1991;Birch等人,Bioprocess Technol.19:251,1990;「Teratocarcinomas and embryonic stem cells:A practical approach」(E.J.Robertson編輯,IRL Press Ltd.1987);「Guide to Techniques in Mouse Development」(P.M.Wasserman等人編輯,Academic Press 1993);「Embryonic Stem Cell Differentiation in vitro」(M.V.Wiles,Meth.Enzymol.225:900,1993);「Properties and uses of Embryonic Stem Cells:Prospects for Application to Human Biology and Gene Therapy」(P.D.Rathjen等人,1993)。
幹細胞的分化在Robertson,Meth.Cell Biol.75:173,1997;和Pedersen,Reprod.Fertil.Dev.10:31,1998中綜述。
10.富集和耗盡策略
在特定實施例中,本發明提供了用於就多能細胞例如iPSC富集細胞群的策略。在一個實施例中,富集提供了用於在相對短的時間內衍生克隆iPSC集落的方法,由此改善iPSC生成的效率。富集可包括分選細胞群,所述細胞群已進行誘導以重編程,以鑑定且獲得表達多能性標記物的細胞,由此獲得富含多能細胞的細胞群。另外的富集方法包括表達分化標記物的細胞或非多能細胞的耗盡,以獲得多能細胞的富集群。在一些實施例中,細胞在誘導重編程後培養約4至30天、約4至24天、約6至22天、或約8至約12天。
在一個實施例中,就多能細胞富集細胞群包括通過使群中的細胞解離且使細胞重懸浮來製備單細胞懸浮液。解離的細胞可重懸浮於任何合適溶液或培養基中,用於維持細胞或執行細胞分選。在特定實施例中,單細胞懸浮液含有GSK3抑制劑、MEK抑制劑和ROCK抑制劑,且缺乏TGFβR抑制劑。在某些實施例中,GSK3抑制劑是CHIR99021,MEK抑制劑是PD0325901,並且ROCK抑制劑是thiazovivin。
在一個特定實施例中,細胞群進行分選,以陽性選擇多能細胞,和/或使群耗盡非重編程或非多能細胞,由此獲得富含多能細胞的細胞群。在一個實施例中,製備單細胞懸浮液,並且隨後單細胞準備用於分選,例如通過使用例如適當抗體就多能性標記物染色。細胞可通過分選細胞的任何合適方法,例如通過磁珠或流式細胞術(FACS)分選進行分選。
細胞可基於各種多能性標記物進行分選,包括SSEA3/4、TRA1-60/81、TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD105、OCT4、NANOG、SOX2、KLF4、SSEA1(小鼠)、CD30、SSEA5、CD90和CD50的表達。在各個實施例中,細胞基於至少兩種、至少三種或至少四種多能性標記物進行分選。在某些實施例中,細胞基於SSEA4的表達進行分選,並且在某些特定實施例中,基於與TRA1-81或TRA1-60組合的SSEA4表達進行分選。在某些實施例中,細胞基於SSEA4、TRA1-81或TRA1-60和CD30表達進行分選。在某些實施例中,細胞最初使用分化細胞的表面標記物包括但不限於CD13、CD26、CD34、CD45、CD31、CD46或CD7耗盡非重編程細胞,並且隨後就多能標記物例如SSEA4、TRA1-81和CD30進行富集。
富含多能細胞的群可置於細胞培養系統例如常規hESC培養基或本發明的細胞培養基中。細胞培養系統可補充有飼養細胞,或任選是無飼養者環境。在一些實施例中,表達多能性標記物的分選細胞置於補充飼養細胞的培養系統中,並且隨後轉移至無飼養者環境。在一個實施例中,細胞培養基是無飼養者環境,且包含GSK3抑制劑、MEK抑制劑和ROCK抑制劑,並且缺乏TGFβ抑制劑。在特定實施例中,GSK3抑制劑是CHIR99021,MEK抑制劑是PD0325901,並且ROCK抑制劑是thiazovivin。在本發明的其他特定實施例中,細胞培養系統是包含MatrigelTM塗布的組織板的無飼養者環境。在一個實施例中,細胞培養系統包含表1中所述的FMM培養基。
富集的細胞群可在本文描述的細胞培養系統中進行培養,以獲得基態iPSC集落,通常在分選後約3至約25天;分選後約5-9天,或分選後約5-7天出現。iPSC集落可進行挑選或分選用於克隆擴增。使用本文考慮的富集策略,細胞群就多能細胞富集至少約3倍、5倍、或10倍或更多倍。
在一些實施例中,使經歷重編程的細胞群或多能細胞群耗盡分化細胞。在一個實施例中,多能細胞群或誘導為重編程的細胞群可耗盡具有分化細胞的細胞表面標記物的細胞。分化細胞的細胞表面標記物的舉例說明性例子包括但不限於CD13、CD26、CD34、CD45、CD31、CD46或CD7。在特定實施例中,CD13用作分化細胞的表面標記物。
在其他實施例中,細胞群進行誘導以分化成所需譜系,且耗盡多能細胞,以獲得分化中細胞或分化細胞的富集群。在一些實施例中,分化細胞群包含已誘導以分化成特異性譜系的細胞群例如ESC或iPSC群。使用上文描述的負細胞分選技術(「淘選」),例如根據磁珠或基於多能性標記物的FAC分選群中的細胞,細胞群可耗盡多能細胞。在一些實施例中,包含分化細胞的細胞群通過FAC使用多能性標記物進行分選,並且獲得耗盡表達多能性標記物的細胞的級分。在其他實施例中,細胞群通過FAC基於分化標記物,例如譜系特異性標記物如CD13、CD26、CD34、CD45、CD31、CD46或CD7進行分選,以獲得耗盡多能性標記物的級分。CD13在本發明的特定實施例中用作分化細胞的表面標記物。
E.用於使細胞重編程的培養平臺
推行各種策略以誘導細胞中的多能性,或增加細胞中的潛能(Takahashi,K.和Yamanaka,S.,Cell 126,663-676(2006);Takahashi等人,Cell 131,861-872(2007);Yu等人,Science 318,1917-1920(2007);Zhou等人,Cell Stem Cell 4,381-384(2009);Kim等人,Cell Stem Cell 4,472-476(2009);Yamanaka等人,2009;Saha,K.,Jaenisch,R.,Cell Stem Cell 5,584-595(2009)),且改善重編程效率(Shi等人,Cell Stem Cell 2,525-528(2008a);Shi等人,Cell Stem Cell 3,568-574(2008b);Huangfu等人,Nat Biotechnol 26,795-797(2008a);Huangfu等人,Nat Biotechnol 26,1269-1275(2008b);Silva等人,Plos Bio 6,e253.doi:10.1371/journal.pbio.0060253(2008);Lyssiotis等人,PNAS 106,8912-8917(2009);Ichida等人,Cell Stem Cell 5,491-503(2009);Maherali,N.,Hochedlinger,K.,Curr Biol 19,1718-1723(2009b);Esteban等人,Cell Stem Cell 6,71-79(2010);Feng等人,Cell Stem Cell 4,301-312(2009))。然而,現有方法仍有待實現用於製造工業或臨床級別多能細胞的高流通量解決方案,所述細胞即具有同質多能性、無顯著的自發分化以及在確定的無異物、飼養細胞培養系統中使用單細胞酶促傳代培養且擴增細胞群的能力的克隆無轉基因多能細胞群。
本文考慮的培養平臺可部分用於生產高級別誘導多能幹細胞(iPSC)。在一個實施例中,非多能細胞重編程為多能性且培養以維持多能性。在另一個實施例中,iPSC培養至基態多能性。
在各個實施例中,培養平臺使無轉基因和/或足跡重編程方法成為可能。本文考慮的培養平臺提供了高度有效的附加體重編程,具有hiPSC生成所需的時間和努力的顯著減少。不希望受任何特定理論束縛,考慮通過在重編程過程中早期阻斷分化跡象且通過特異性途徑(MEK、ERK、TGFβ和ROCK)的小分子抑制促進間充質至上皮轉變(MET),hiPSC生成的效率使用附加型載體在FF和單細胞培養系統中得到顯著改善。
在一個實施例中,培養平臺包括使一個或多個非多能細胞重編程為多能狀態,包括增加細胞中的內源OCT4表達。細胞中的內源OCT4表達可通過引入OCT4表達的一種或多種多核苷酸、多肽或小分子誘導物得到增加。在一個實施例中,將編碼OCT4的多核苷酸或OCT4多肽引入細胞內足以誘導OCT4在細胞中的內源表達。
在一個實施例中,培養平臺包括使一個或多個非多能細胞重編程,包括將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC和SV40LT。在另一個實施例中,培養平臺包括使一個或多個非多能細胞重編程,包括將選自下述的一種或多種多肽引入一個或多個非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1。
在一個實施例中,培養平臺包括使一個或多個非多能細胞重編程,包括將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸引入一個或多個非多能細胞內:OCT4、NANOG、ESRRB、ECAT1和UTF1。在另一個實施例中,培養平臺包括使一個或多個非多能細胞重編程,包括將選自下述的一種或多種多肽引入一個或多個非多能細胞內:OCT4、NANOG、ESRRB、ECAT1和UTF1。
如本文使用的,在特定實施例中,術語「引入」指包括使細胞與多核苷酸、多肽或小分子接觸的過程。引入步驟還可包括將多核苷酸或多肽顯微注射到細胞內,使用脂質體以將多核苷酸或多肽遞送到細胞內,或使多核苷酸或多肽與細胞可滲透部分融合以將其引入細胞內。
在特定實施例中,編碼選自下述的重編程因子中的一種或多種的1、2、3、4、5個或更多個拷貝的一種或多種多核苷酸可引入非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1,以使細胞重編程。引入細胞內的每種重編程因子的拷貝數在適合於實現如本文考慮的基態多能性的任何組合中可以是相同或不同的。
在一個實施例中,將編碼選自下述的重編程因子中的一種或多種的1、2、3、4、5個或更多個拷貝的一種或多種多核苷酸引入非多能細胞內:OCT4、SOX2和NANOG。
在一個實施例中,將編碼OCT4、SOX2和NANOG各自的1、2、3、4、5個或更多個拷貝的一種或多種多核苷酸引入非多能細胞內。
在一個實施例中,將編碼OCT4和SOX2各自的1、2、3、4、5個或更多個拷貝的一種或多種多核苷酸引入非多能細胞內。
在一個實施例中,將編碼OCT4的1、2、3、4、5個或更多個拷貝的一種或多種多核苷酸引入非多能細胞內。
在一個實施例中,將編碼OCT4的2個拷貝、SOX2的2個拷貝和NANOG的2個拷貝的一種或多種多核苷酸引入非多能細胞內,其中SV40LT任選被引入非多能細胞內。
在另一個實施例中,將編碼OCT4的3個拷貝、SOX2的2個拷貝、NANOG的一個拷貝和UTF1的一個拷貝的一種或多種多核苷酸引入非多能細胞內。
在各個舉例說明性實施例中,包括使非多能細胞重編程的培養平臺包括引入編碼OCT4的多核苷酸的1至5個拷貝;編碼SOX2的多核苷酸的1至3個拷貝,以及任選地,編碼NANOG的多核苷酸的1至2個拷貝。多核苷酸可作為一種或多種更大多核苷酸的任何組合引入細胞內。在一個非限制性例子中,將編碼OCT4的1至4個拷貝、SOX2的1或2個拷貝和NANOG的1個拷貝的一種或多種多核苷酸引入非多能細胞內。在另一個非限制性例子中,使非多能細胞重編程為多能狀態包括將編碼OCT4的2個拷貝的第一多核苷酸、編碼OCT4的1個拷貝和SOX2的1個拷貝的第二多核苷酸;以及編碼OCT4的1個拷貝、SOX2的1個拷貝和NANOG的1個拷貝的第三多核苷酸引入非多能細胞內。在進一步的非限制性例子中,使一個或多個非多能細胞重編程包括將編碼OCT4的2個拷貝的第一多核苷酸以及編碼OCT4的1個拷貝、SOX2的1個拷貝和NANOG的1個拷貝的第二多核苷酸引入一個或多個非多能細胞內。在再進一步的非限制性例子中,將編碼OCT4的2個拷貝的第一多核苷酸以及編碼OCT4的1個拷貝和SOX2的1個拷貝的第二多核苷酸引入一個或多個非多能細胞內,以產生多能細胞。
在一個實施例中,將包括包含本文考慮的重編程因子的任何數目和組合的多核苷酸的單一載體引入非多能細胞內,且足以使細胞重編程為多能狀態。
在一個實施例中,將包括包含本文考慮的重編程因子的任何數目和組合的1、2、3、4、5種或更多種多核苷酸的一種或多種載體引入非多能細胞內,且足以使細胞重編程為多能狀態。
在一個優選實施例中,用於使非體細胞重編程的包含本文考慮的一種或多種多核苷酸的一種或多種載體用於將一種或多種多核苷酸引入細胞內,且足以使細胞重編程。
在最優選的實施例中,用於使非體細胞重編程的包含本文考慮的一種或多種多核苷酸的一種或多種附加型載體用於將一種或多種多核苷酸引入細胞內,且足以使細胞重編程。展示減少的自發分化和/或基態的多能細胞可用如本文考慮的附加型載體進行製造,並且隨後培養直至載體喪失,以獲得展示減少的自發分化和/或基態的多能細胞,所述多能細胞不包含編碼重編程因子的外源核酸。
還考慮當包含其的多核苷酸或載體包括編碼至少兩種重編程因子或重編程因子的至少兩個拷貝的多核苷酸時,多核苷酸包含在如本文考慮的重編程因子各自之間的IRES序列或編碼自切割多肽序列的多核苷酸。
在一些方面,在將重編程因子多核苷酸引入非多能細胞內之後,通過就一種或多種重編程因子多核苷酸的異位表達進行選擇,增加使非多能細胞重編程的效率。此類選擇可例如通過下述發生:使重編程因子多核苷酸中的一種或多種與可選標記物連接,將重編程因子多核苷酸和可選標記物引入非多能細胞內,並且選擇表達可選標記物的那些細胞,其中所述選擇鑑定相對於缺乏標記物及其相關的重編程因子多核苷酸的表達的細胞具有增加的重編程效率的細胞。本領域技術人員應了解在特定實施例中,可使用鑑定通過非多能細胞的所引入重編程多核苷酸表達的任何可選標記物。
此類可選標記物的一個非限制性例子包括但不限於抗生素抗性基因例如嘌呤黴素抗性。可選標記物可與下述重編程因子多核苷酸中的一種或多種連接:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1。在一些實施例中,重編程因子多核苷酸的特異性組合作為多順反子載體引入,其中可選標記物連接至重編程因子多核苷酸的特異性組合。重編程因子多核苷酸的特異性組合可編碼本文公開的重編程因子多核苷酸的兩個或更多個拷貝。
在一個非限制性實施例中,多順反子載體編碼與可選標記物例如編碼嘌呤黴素抗性的基因連接的OCT4多核苷酸的兩個或更多個拷貝。
在一些方面,除一種或多種分開的重編程因子多核苷酸之外,還將編碼一種或多種重編程因子多核苷酸的多順反子載體和可選標記物引入非多能細胞內,其中就表達可選標記物的細胞進行選擇產生具有比缺乏可選標記物表達的細胞更大的重編程效率的細胞群。
在該選擇過程的一個非限制性例子中,除編碼與嘌呤黴素抗性基因連接的OCT4的兩個或更多個拷貝的多順反子載體之外,還將OCT4、NANOG和SOX2多核苷酸引入非多能細胞內。表達可選標記物的非多能細胞的後續選擇鑑定相對於不表達可選標記物的非多能細胞,具有更大的重編程效率的非多能細胞。所選細胞可具有至少5%、至少10%、至少15%、至少20%、至少30%或至少40%的重編程效率。
小分子通常包括在特定優選實施例的重編程步驟中。不希望受任何特定理論束縛,考慮各種分化途徑的小分子抑制劑的包括增加重編程效率和動力學。相應地,在特定實施例中,包含使非多能細胞重編程的培養平臺包括如本文考慮的將一種或多種重編程因子引入細胞內,並且使細胞與GSK3抑制劑;MEK抑制劑;TGFβR抑制劑和ROCK抑制劑接觸。
重編程效率的改善可通過下述進行測量:(1)多能細胞重編程和生成所需的時間的減少(例如與不含小分子的相似或相同過程相比較,通過使生成多能細胞的時間縮短至少一天),或可替代地或與(2)組合,(2)通過特定過程生成的多能細胞數目的增加(例如與不含小分子的相似或相同過程相比較,給定時間段中重編程的細胞數目增加至少10%、30%、50%、100%、200%、500%等)。在一些實施例中,觀察到重編程效率的2倍至20倍改善。在一些實施例中,重編程效率改善超過20倍。在一些實施例中,觀察到超過不含小分子重編程試劑的方法超過100倍的效率改善(例如生成的多能細胞數目的超過100倍增加)。
在一個實施例中,通過如本文考慮的將一種或多種重編程因子引入細胞內,且使細胞與GSK3抑制劑;MEK抑制劑;和TGFβR抑制劑,以及任選的ROCK抑制劑接觸,本文考慮的培養平臺包括使非多能細胞重編程。
在一個優選實施例中,通過如本文考慮的將一種或多種重編程因子引入細胞內,且使細胞與GSK3抑制劑;MEK抑制劑;TGFβR抑制劑以及ROCK抑制劑接觸,本文考慮的培養平臺包括使非多能細胞重編程。
在一個更優選的實施例中,通過如本文考慮的將一種或多種重編程因子引入細胞內,且使細胞與GSK3抑制劑;MEK抑制劑;TGFβR抑制劑以及ROCK抑制劑接觸,本文考慮的培養平臺包括使非多能細胞重編程,其中所述ROCK抑制劑是Thiazovivin。
然而,為了使多能細胞在無飼養細胞和酶促傳代的培養系統中的長期培養成為可能,伴隨減少的自發分化或無顯著的自發分化,或者為了誘導和/或維持基態多能性,iPSC需要在包含GSK-3抑制劑、MEK抑制劑和任選的Rho激酶(ROCK)抑制劑的細胞培養基中的後續培養,其中所述細胞培養基不包含或缺乏TGFβ/激活素信號傳導途徑的抑制劑,包括TGFβ受體(TGFβR)抑制劑和ALK5抑制劑,如本文考慮的。不希望受任何特定理論束縛,考慮多能細胞與TGFβR/ALK5抑制劑一起長期培養導致培養的無轉基因iPSC的自發分化和基態多能性的最終喪失。
在各個實施例中,兩步培養平臺用於使體細胞穩定重編程,以實現培養中減少的自發分化,包括基態多能性。在某些實施例中,非多能細胞通過本領域公開的任何合適方法進行重編程,並且隨後,通過在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養細胞,來培養重編程的體細胞以實現培養中減少的自發分化,其中所述培養基缺乏TGFβR/ALK5抑制劑。在一些實施例中,培養重編程的體細胞以提供基態多能細胞。
在特定實施例中,非多能細胞通過本文公開的方法進行重編程,並且隨後,通過在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養細胞,將重編程的體細胞培養至穩定的基態多能性,其中所述培養基缺乏TGFβR/ALK5抑制劑。
在一些實施例中,通過引入一種或多種重編程因子,並且在包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和TGFβR/ALK5抑制劑的培養基中培養細胞,使非多能細胞重編程,並且隨後,通過在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養細胞,來培養重編程的體細胞以提供具有減少的自發分化的細胞,其中所述培養基缺乏TGFβR/ALK5抑制劑。
在一些實施例中,通過引入一種或多種重編程因子,並且在包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和TGFβR/ALK5抑制劑的培養基中培養細胞,使非多能細胞重編程,並且隨後,通過在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養細胞,將重編程的體細胞培養至穩定的基態多能性,其中所述培養基缺乏TGFβR/ALK5抑制劑。
在優選實施例中,通過引入一種或多種重編程因子,並且在包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和TGFβR/ALK5抑制劑的培養基中培養細胞,使非多能細胞重編程,並且隨後,通過在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養細胞,將重編程的體細胞培養至穩定的基態多能性,其中所述培養基缺乏TGFβR/ALK5抑制劑,並且其中不存在重編程轉基因的顯著殘留表達。
在一個實施例中,通過引入如本文其他地方公開的選自下述的一種或多種重編程因子:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1,並且在包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和TGFβR/ALK5抑制劑的培養基中培養細胞,使非多能細胞重編程,並且隨後,在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養重編程的體細胞,其中所述培養基缺乏TGFβR/ALK5抑制劑。
在一個優選實施例中,通過引入如本文其他地方公開的選自下述的一種或多種重編程因子:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1,並且在包含GSK-3抑制劑、MEK抑制劑、Rho激酶(ROCK)抑制劑和TGFβR/ALK5抑制劑的培養基中培養細胞,使非多能細胞重編程,並且隨後,在包含GSK-3抑制劑、MEK抑制劑和Rho激酶(ROCK)抑制劑的培養基中培養重編程的體細胞,其中所述ROCK抑制劑是Thiazovivin,其中所述培養基缺乏TGFβR/ALK5抑制劑。
在各個實施例中,本發明提供了使用本文考慮的培養平臺,製造具有減少的自發分化的多能細胞和/或基態誘導多能幹細胞(iPSC)的方法。
在特定實施例中,使用包含一個或多個非多能細胞或部分多能幹細胞的原材料,並且在不包含TGFβR抑制劑的培養基中培養一個或多個多能細胞或部分多能幹細胞,製造具有減少的自發分化的多能細胞和/或基態誘導多能幹細胞(iPSC)。原材料可獲得或製備。例如,非多能幹細胞或部分多能幹細胞可由商業廠商或其他源提供,或者可從頭獲得:非多能細胞還可從組織或器官中分離;並且部分多能細胞還可通過使體細胞或成人幹細胞重編程來生成。在一些實施例中,使用本文描述的培養基和平臺,多能胚胎幹細胞或通過體細胞核轉移獲得的多能細胞可進行誘導,以實現基態多能性。
在特定實施例中,一個或多個IPSC群可包含重編程的體細胞或重編程的成人幹細胞。在特定實施例中,可通過任何已知方法通過執行方法或獲得通過方法生成的IPSC來生成IPSC。
生成IPSC的示例性方法包括但不限於:增加非多能細胞中的內源OCT4表達;將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸,任選以一個或多個拷貝引入一個或多個非多能細胞內:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1;或將編碼選自下述的一種或多種重編程因子的一種或多種多核苷酸,任選以一個或多個拷貝引入一個或多個非多能細胞內:OCT4、SOX2和NANOG。生成IPSC的方法還可包括使一個或多個非多能細胞或部分多能細胞與GSK3抑制劑;MEK抑制劑;和TGFβR抑制劑,以及任選的ROCK抑制劑接觸,以產生一個或多個iPSC。
在某些實施例中,細胞培養基包含GSK3抑制劑;MEK抑制劑;和ROCK抑制劑。
在優選實施例中,細胞培養基包含GSK3抑制劑;MEK抑制劑;和ROCK抑制劑,其中所述ROCK抑制劑是Thiazovivin。
在特定實施例中,在細胞培養基中培養一個或多個多能細胞例如IPSC維持或誘導基態多能性、活力、正常核型、基因組穩定性和減少速率的自發分化,其可維持至少5次傳代、至少10次傳代、至少50次傳代、至少100次傳代或更多次傳代,包括任何中間數目的傳代。
F.表徵多能細胞
使用本文考慮的培養平臺製造的多能細胞還可包括多能細胞產物,包括例如基態多能細胞或具有減少的自發分化的多能細胞的選擇或驗證。多能細胞可在重編程且隨後用本文考慮的組合物和方法進行培養之後,或如果多能細胞未重編程,在多能細胞轉變至本文考慮的培養方法之後,進行選擇和/或驗證。細胞的多能性可基於與多能性相關的有關和可檢測的形態、分子和/或生物化學變化進行表徵和/或選擇。
在評價細胞潛能中可分開或組合監控的細胞多能性的具體特徵包括但不限於基因表達、甲基化以及體內和體外特徵,例如:i)圓形的多能幹細胞形態;ii)多能幹細胞標記物包括SSEA3/4(人多能幹細胞);TRA1-60/81;TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD105、OCT4、NANOG、SOX2、CD30、SSEA5、CD90和/或CD50和前述的組合的表達;iii)多能幹細胞的畸胎瘤形成;iv)胚狀體形成和體外三系分化;和v)失活的X染色體復活。在某些實施例中,上述特徵中的任一個的亞集用於監控細胞潛能。在一個實施例中,多能細胞的特徵在於具有圓形集落形態,SSEA4、TRA1-81和OCT4的表達,以及形成胚狀體和畸胎瘤的能力。
在另一個實施例中,在體外培養中具有減少的自發分化的多能細胞可通過基因表達標誌進行鑑定,所述基因表達標誌包括與在TGFβR抑制劑的存在下培養的多能細胞相比較,下述分化標記物基因中的一種或多種表達中的至少約10%、20%、25%、30%、40%、50%、60%、70%、80%、90%減少:FOXA2、FGF5、SOX17、XIST、NODAL、COL3A1、OTX2、DUSP6、EOMES、NR2F2、NR0B1、CXCR4、CYP2B6、GATA3、GATA4、ERBB4、GATA6、HOXC6、INHA、SMAD6、RORA、NIPBL、TNFSF11、CDH11、ZIC4、GAL、SOX3、PITX2、APOA2、CXCL5、CER1、FOXQ1、MLL5、DPP10、GSC、PCDH10、CTCFL、PCDH20、TSHZ1、MEGF10、MYC、DKK1、BMP2、LEFTY2、HES1、CDX2、GNAS、EGR1、COL3A1、TCF4、HEPH、KDR、TOX、FOXA1、LCK、PCDH7、CD1D FOXG1、LEFTY1、TUJ1、T基因(Brachyury)和ZIC1。
在一個實施例中,具有減少的自發分化的多能細胞的特徵在於一種或多種分化標記物基因的減少表達,所述基因包括但不限於:T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2和TUJ1。在特定實施例中,具有減少的自發分化的多能細胞可通過基因表達標誌進行鑑定,所述基因表達標誌包括與在TGFβR抑制劑的存在下培養的多能細胞相比較,一種或多種分化標記物基因(例如T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2、TUJ1)的表達中的至少約10%、20%、25%、30%、40%、50%、60%、70%、80%、90%減少。[在另一個特定實施方案中,具有減少的自發分化的多能細胞可通過基因表達標誌進行鑑定,所述基因表達標誌包括一種或多種分化標記物基因(例如T基因、CXCR4、NODAL、GATA4、SOX17、FOXA2、OTX2、TUJ1)的表達中的至少約10%、20%、25%、30%、40%、50%、60%、70%、80%、90%減少。
在特定實施例中,基態多能細胞具有顯著阻遏的Xist表達以及分化細胞的早期標記物例如Foxa2、Sox17和Brachyury的表達,而常規培養的多能細胞僅顯示Xist表達的適度阻遏以及早期分化標記物的顯著表達。
在特定實施例中,基態多能細胞保留基態多能性的特徵共多個細胞傳代,例如至少1、3、5、7、10、15、20次或更多次傳代。
G.多核苷酸
在各個舉例說明性實施例中,本發明部分考慮了本文考慮的多核苷酸、編碼多肽和融合多肽的多核苷酸、以及包含其的組合物。在各個其他舉例說明性實施例中,本發明部分考慮了用編碼至少一種重編程因子的一個或多個拷貝的多核苷酸使非多能細胞重編程。用於與本文描述的培養平臺一起使用的重編程因子包括但不限於:OCT4、SOX2、NANOG、KLF4、LIN28、C-MYC、SV40LT、hTERT、SALL4、GLIS、ESRRB、DPPA2、ECAT1、SOX1、SOX3、KLF2、KLF5、L-MYC、N-MYC、LRH1和UTF1。在優選實施例中,多核苷酸包含如本文所示的重編程因子的序列。
如本文使用的,術語「基因」可指包含增強子、啟動子、內含子、外顯子等等的多核苷酸序列。在特定實施例中,術語「基因」指編碼多肽的多核苷酸序列,與多核苷酸序列是否等同於編碼多肽的基因組序列無關。
如本文使用的,「經分離的多核苷酸」指已從序列中純化的多核苷酸,所述序列在天然存在的狀態中側接其,例如已從與片段通常鄰近的序列中去除的DNA片段。在特定實施例中,「經分離的多核苷酸」指在自然界中不存在且已通過人工製備的互補DNA(cDNA)、重組DNA或其他多核苷酸。
在特定實施例中,一種或多種多核苷酸可以任何合適次序排列在更大的多核苷酸例如載體內。在優選實施例中,載體是附加型載體。
與其編碼序列長度無關,本文考慮的多核苷酸可與其他DNA序列組合,例如表達控制序列、啟動子和/或增強子、非翻譯區(UTR)、Kozak序列、多腺苷酸化信號、另外的限制性酶位點、多克隆位點、內部核糖體進入位點(IRES)、重組酶識別位點(例如LoxP、FRT和Att位點)、終止密碼子、轉錄終止信號、和編碼自切割多肽的多核苷酸、附加表位,如本文其他地方公開的或如本領域已知的,使得其總體長度可相當大地改變。因此考慮可採用幾乎任何長度的多核苷酸片段,其中總長度優選受製備的容易和在預期重組DNA方案中的使用限制。
多核苷酸可使用本領域已知和可獲得的各種充分確定的技術中的任一種進行製備、操縱和/或表達。為了表達所需多肽,編碼多肽的核苷酸序列可插入適當載體內。載體的例子是質粒、自主複製序列和可易位元件。另外的示例性載體包括但不限於質粒,噬菌粒,粘粒,人工染色體如酵母人工染色體(YAC)、細菌人工染色體(BAC)、或P1衍生的人工染色體(PAC),噬菌體如λ噬菌體或M13噬菌體,以及動物病毒。可用作載體的動物病毒類別的例子包括但不限於逆轉錄病毒(包括慢病毒)、腺病毒、腺相關病毒、皰疹病毒(例如單純皰疹病毒)、痘病毒、杆狀病毒、乳頭狀瘤病毒和乳多空病毒(例如SV40)。表達載體的例子是用於在哺乳動物細胞中表達的pClneo載體(Promega);用於慢病毒介導的基因轉移和在哺乳動物細胞中表達的pLenti4/V5-DESTTM、pLenti6/V5-DESTTM和pLenti6.2/V5-GW/lacZ(Invitrogen)。在特定實施例中,本文公開的多肽的編碼序列可連接到此類表達載體內,用於在哺乳動物細胞中表達多肽。
在特定實施例中,載體是附加型載體或染色體外維持的載體。如本文使用的,術語「附加型」指無需整合到宿主的染色體DNA內能夠複製且不具有從分裂中宿主細胞的逐步喪失的載體,還意指所述載體在染色體外或附加型複製。載體進行改造,以容納來自嗜淋巴皰疹病毒或γ皰疹病毒、腺病毒、SV40、牛乳頭狀瘤病毒或酵母的編碼DNA複製起點或「ori」的序列,具體地嗜淋巴皰疹病毒或γ皰疹病毒的複製起點,對應於EBV的oriP。在一個特定方面,嗜淋巴皰疹病毒可以是EB病毒(EBV)、卡波濟氏肉瘤皰疹病毒(KSHV)、松鼠猴皰疹病毒(HS)或馬立克氏病病毒(MDV)。EB病毒(EBV)和卡波濟氏肉瘤皰疹病毒(KSHV)也是γ皰疹病毒的例子。通常,宿主細胞包含活化複製的病毒複製反式激活蛋白質。
表達載體中存在的「表達控制序列」、「控制元件」或「調節序列」是載體的那些非翻譯區—複製起點、選擇盒、啟動子、增強子、翻譯起始信號(Shine Dalgarno序列或Kozak序列)、內含子、多聚腺苷酸化序列,5'和3'非編碼區—所述非翻譯區與宿主細胞蛋白質相互作用,以進行轉錄和翻譯。此類元件可在其強度和特異性上不同。取決於所利用的載體系統和宿主,可使用任何數目的合適轉錄和翻譯元件,包括遍在啟動子和誘導型啟動子。
術語「可操作地連接」指其中所述組分處於允許其以其預期方式起作用的關係中的並排。在一個實施例中,該術語指表達控制序列(例如啟動子和/或增強子)和第二多核苷酸序列之間的功能連接,其中表達控制序列指導對應於第二序列的核酸的轉錄。
適用於本發明的特定實施例中的舉例說明性遍在表達控制序列包括但不限於巨細胞病毒(CMV)立即早期啟動子,猿猴病毒40(SV40)(例如早期或晚期),莫洛尼鼠白血病病毒(MoMLV)LTR啟動子,Rous肉瘤病毒(RSV)LTR,單純皰疹病毒(HSV)(胸苷激酶)啟動子,來自牛痘病毒的H5、P7.5和P11啟動子,延伸因子1-α(EF1a)啟動子,早期生長應答1(EGR1),鐵蛋白H(FerH),鐵蛋白L(FerL),甘油醛3-磷酸脫氫酶(GAPDH),真核翻譯起始因子4A1(EIF4A1),熱休克70kDa蛋白5(HSPA5),熱休克蛋白90kDaβ成員1(HSP90B1),熱休克蛋白70kDa(HSP70),β-驅動蛋白(β-KIN),人ROSA 26基因座(Irions等人,Nature Biotechnology 25,1477-1482(2007)),泛素C啟動子(UBC),磷酸甘油酸激酶-1(PGK)啟動子,巨細胞病毒增強子/雞β-肌動蛋白(CAG)啟動子和β-肌動蛋白啟動子。
誘導型啟動子/系統的舉例說明性例子包括但不限於甾類誘導型啟動子,例如用於編碼糖皮質激素或雌激素受體的基因的啟動子(可通過用相應激素處理誘導)、金屬硫蛋白啟動子(可通過用各種重金屬處理誘導)、MX-1啟動子(可通過幹擾素誘導)、「基因開關」米非司酮可調節系統(Sirin等人,2003,Gene,323:67)、cumate誘導型基因開關(WO 2002/088346)、四環素依賴性調節系統等。
條件表達還可通過使用位點特異性DNA重組酶來實現。根據本發明的某些實施例,多核苷酸包含關於通過位點特異性重組酶介導的重組的至少一個(通常為兩個)位點。如本文使用的,術語「重組酶」或「位點特異性重組酶」包括切除或整合蛋白質、酶、輔因子或相關蛋白質,所述相關蛋白質涉及牽涉一個或多個重組位點(例如二、三、四、五、六、七、八、九、十個或更多個)的重組反應,所述相關蛋白質可以是野生型蛋白質(參見Landy,Current Opinion in Biotechnology 3:699-707(1993)),或其突變體、衍生物(例如含有重組蛋白質序列或其片段的融合蛋白)、片段和變體。適用於本發明的特定實施例中的重組酶的舉例說明性例子包括但不限於:Cre、Int、IHF、Xis、Flp、Fis、Hin、Gin、ΦC31、Cin、Tn3解離酶、TndX、XerC、XerD、TnpX、Hjc、Gin、SpCCE1和ParA。
在特定實施例中,本文考慮的多核苷酸包括編碼一種或多種多肽的一種或多種多核苷酸。在特定實施例中,為了實現多重多肽各自的有效翻譯,多核苷酸序列可通過一種或多種IRES序列或編碼自切割多肽的多核苷酸序列分開。如本文使用的,「內部核糖體進入位點」或「IRES」指促進對順反子(蛋白質編碼區)的起始密碼子例如ATG的直接內部核糖體進入的元件,由此導致基因的帽不依賴性翻譯。參見例如Jackson等人,1990.Trends Biochem Sci15(12):477-83)以及Jackson和Kaminski.1995.RNA 1(10):985-1000。由本領域技術人員一般採用的IRES的例子包括美國專利號6,692,736中描述的那些。本領域已知的「IRES」的進一步例子包括但不限於可得自小核糖核酸病毒的IRES(Jackson等人,1990)。
H.多肽
本發明部分考慮了包含多肽、融合多肽和表達多肽的載體的組合物。在優選實施例中,多肽包含本文闡述的胺基酸序列。「多肽」、「多肽片段」、「肽」和「蛋白質」可互換使用,除非有相反的說明,且根據常規含義,即作為胺基酸的序列。在一個實施例中,「多肽」包括融合多肽及其他變體。多肽可使用各種眾所周知的重組和/或合成技術中的任一種進行製備。多肽並不限於特定長度,例如它們可包含全長蛋白質序列、全長蛋白質的片段或融合蛋白,並且可包括多肽的翻譯後修飾,例如糖基化、乙醯化、磷酸化等等,以及本領域已知的其他修飾,天然存在和非天然存在的兩者。
如本文使用的,「經分離的肽」或「經分離的多肽」等等指肽或多肽分子從細胞環境和與細胞的其他組分結合中的體外分離和/或純化,即它不與體內物質顯著結合。
在一個實施例中,當需要兩種或更多種多肽的表達時,編碼其的多核苷酸序列可通過如本文其他地方討論的IRES序列分開。在另一個實施例中,兩種或更多種多肽可作為融合蛋白表達,所述融合蛋白包含在本文描述的多肽結構域各自之間的多肽切割信號。另外,多肽位點可置於任何接頭肽序列內。示例性多肽切割信號包括多肽切割識別位點,例如蛋白酶切割位點、核酸酶切割位點(例如罕見限制性酶識別位點、自切割核酶識別位點)和自切割病毒寡肽(參見deFelipe和Ryan,2004.Traffic,5(8);616-26)。
合適的蛋白酶切割位點和自切割肽是技術人員已知的(參見例如Ryan等人,1997.J.Gener.Virol.78,699-722;Scymczak等人(2004)Nature Biotech.5,589-594)。示例性蛋白酶切割位點包括但不限於馬鈴薯Y病毒NIa蛋白酶(例如菸草蝕紋病毒蛋白酶)、馬鈴薯Y病毒HC蛋白酶、馬鈴薯Y病毒P1(P35)蛋白酶、byovirus NIa蛋白酶,byovirus RNA-2編碼的蛋白酶,口蹄疫病毒L蛋白酶、腸道病毒2A蛋白酶、鼻病毒2A蛋白酶、picorna 3C蛋白酶、豇豆花葉病毒24K蛋白酶、線蟲傳多面體病毒24K蛋白酶、RTSV(水稻東格魯球狀病毒)3C樣蛋白酶、PYVF(歐防風黃點病毒)3C樣蛋白酶、肝素、凝血酶、因子Xa和腸激酶的切割位點。由於其高切割嚴格性,TEV(菸草蝕紋病毒)蛋白酶切割位點在一個實施例中是優選的,例如EXXYXQ(G/S)(SEQ ID NO:29),例如ENLYFQG(SEQ ID NO:30)和ENLYFQS(SEQ ID NO:31),其中X代表任何胺基酸(通過TEV的切割在Q和G或Q和S之間發生)。
在某些實施例中,自切割多肽位點包含2A或2A樣位點、序列或結構域(Donnelly等人,2001.J.Gen.Virol.82:1027-1041)。在一個特定實施例中,病毒2A肽是口蹄疫病毒2A肽、馬鈴薯Y病毒2A肽或心病毒2A肽。
在一個實施例中,病毒2A肽選自:口蹄疫病毒(FMDV)2A肽、馬鼻炎A病毒(ERAV)2A肽、明脈扁刺蛾β四體病毒(Thosea asigna virus)(TAV)2A肽、豬捷申病毒-1(PTV-1)2A肽、泰勒病毒2A肽和腦心肌炎病毒2A肽。
表2:示例性2A位點包括下述序列:
在優選實施例中,編碼一種或多種重編程因子多肽的載體包含在重編程因子各自之間的相同或不同蛋白酶切割位點中的一個或多個。
本說明書中引用的所有出版物、專利申請和頒發專利以引用的方式併入本文,如同每個個別出版物、專利申請或頒發專利特別且各別指出以引用的方式併入。
儘管本發明已通過舉例說明和例子略微詳細地進行描述用於明確理解的目的,但對於本領域普通技術人員顯而易見的是,根據本發明的教導,可對其作出某些改變和修飾,而不背離所附權利要求的精神或範圍。提供下述實例僅作為例子而不是限制。本領域技術人員容易認識到可改變或修飾以獲得大體上相似的結果的各種非關鍵參數。
實例
本文公開的實例鑑定了使用在階段特異性培養基組合物中的小分子途徑抑制劑,用於hiPSC的快速、平行生成、選擇和擴增的平臺。該平臺支持在完全無飼養者環境中使用最低限度重編程因子的有效和加速的附加體重編程。所得到的hiPSC不含轉基因,容易作為單細胞培養且擴增,同時維持同質和基因組穩定的多能群。在實例中考慮的培養基組合物中生成或維持的hiPSC顯示出與基態多能性相關的特性,並且代表用於製造均勻的工業或臨床級別hiPSC的穩健高流通量系統。
實例1
用於iPSC的長期維持和擴增的培養基平臺的鑑定
概述
在SMC4補充的培養中的大多數慢病毒衍生的hiPSC系維持未分化細胞的同質群;然而,在系亞集中的轉基因重編程因子的沉默展示在擴增培養中不同程度的自發分化(圖1A和B)。因此,評價各種細胞培養組分,以便鑑定與殘留轉基因表達無關,在連續FF培養和單細胞酶促傳代期間維持多能性的條件。靶向重編程和維持過程的不同階段時的獨特途徑的多階段培養系統鑑定為hiPSC生成的有效和穩健方法。
結果
在長期維持期間TGFβ途徑的抑制鑑定為具有沉默的轉基因表達的hiPSC系的自發分化中的顯著因子(圖1C)。將發現經歷自發分化的iPSC細胞系之一轉變為在新培養基製劑即命運維持培養基(FMM)中的培養(表1)。自發分化被消除,並且在2-3代內建立SSEA4/TRA1-81陽性細胞的同質群(圖1A)。
還比較了在常規培養(在MEF飼養細胞上的hESC培養基)、在FF培養中補充SMC4的培養基或在FF培養中新近配製的FMM(圖1D)中的OCT4/KLF4/SOX2(OKS)慢病毒重編程。在慢病毒重編程誘導後十七天,SSEA4/TRA1-81陽性細胞通過FAC加以選擇,並且在SMC4或FMM中再鋪平板用於比較(圖1D)。SMC4改善重編程的動力學,並且在誘導後第17天時導致比用FMM重編程顯著更多的SSEA4/TRA1-81陽性細胞(2.72%相對於FMM的0.76%和常規培養的0.10%;圖1D)。
在初始分選後,細胞在其分別條件下維持10天,隨後為第二SSEA4/TRA1-81陽性流式細胞術選擇(圖1D)。培養維持另外9天(總共轉染後36天),並且基於OCT4和NANOG共表達就未分化集落進行評分(圖1D和1E)。在SMC4中的初始重編程隨後轉變為FMM的組合最終導致相對於在SMC4中的連續維持,更多OCT4/NANOG陽性集落和顯著減少數目的OCT4/NANOG陰性集落(圖1D和1E)。儘管OCT4/NANOG陽性集落在FMM中專一維持的培養物中檢測到,但集落數目和大小看起來不如階段特異性培養基方法。
這些結果顯示靶向重編程和維持過程的不同階段時的獨特途徑的新型多階段培養系統導致高質量hiPSC的有效製造。
實例2
用於製造在單細胞傳代和FF形式中的無轉基因hiPSC的平臺
概述
使用附加型載體系統的非整合重編程方法的效率極低(1%)(圖8A和8B)。在第14天時,將由FRM支持的重編程培養物拆分到FRM或FMM培養基內。在第21天時,FACS用於鑑定培養中的SSEA4/TRA1-81/CD30陽性細胞(圖8C)。FRM維持的培養物含有分化細胞和未分化細胞兩者,而FMM培養物主要含有未分化細胞(圖8D)。
這種方法的流通量和穩健性用成纖維細胞和CD34+細胞進行測試,所述CD34+細胞由來自不同年齡、性別和種族性的供體的最低體積的臍帶血擴增(圖9A和9B)。體細胞重編程如圖2A中概述的使用附加體基因組合集合OSNKLMT進行誘導,並且96孔板流式細胞術在第16和21天之間就個別hiPSC進行分選(圖2B)。對於測試的大多數系觀察到SSEA4/TRA1-81/CD30陽性細胞的大群。與使用常規培養基和飼養細胞的平行重編程實驗相比較,FRM和FMM培養基系統導致hiPSC克隆數目的顯著增加(對於FTC007成纖維細胞系,FRM/FMM中的8.55%相對於常規培養中的0.02%;圖2B和2C)。對於每個體細胞系可見平均起來22個克隆hiPSC/96孔板(圖2B、10A和10B),所述體細胞系包括先前已觀察到對用SMC4培養基的慢病毒重編程抗拒的成纖維細胞FTC008。集落隨後通過分析細胞內和表面標記物表達以及關於NANOG的直接qRTPCR證實為真正的hiPSC克隆(圖2D、2E和10C)。使用96孔分選和選擇過程的重編程效率也得到增加(圖2E)。用確定的表面塗布玻連蛋白也觀察到相似的重編程效率(圖10D)。
這些數據顯示當應用於附加體重編程時,平臺是穩健和可重現的,並且允許以高流通量形式平行執行多重重編程實驗,伴隨最低限度的努力且不妥協iPSC終末產物的質量。
實例3
無轉基因hiPSC系在FMM中的長期傳代和擴增
概述
使用在FF培養中作為單細胞擴增的來自實例2的hiPSC克隆,研究使用FRM和FMM的多階段培養基平臺的hiPSC的長期傳代和擴增(圖3A和3B)。
結果
根據實例2重編程的hiPSC繫到第4-7代時喪失附加型DNA,並且因此不依賴於基於轉基因的重編程因子而多能(圖3C)。hiPSC系維持對於SSEA4、TRA1-81、OCT4和NANOG陽性的未分化細胞的同質群。此外,這些系在不存在任何清除策略的情況下維持多能特徵(圖3F),所述清除策略常用於多能培養中(圖3D和3E)。當在常規的單細胞傳代培養期間,Matrigel替換為確定的表面塗布玻連蛋白時,觀察到均勻hiPSC培養物的相似擴增(圖10E-10G)。
基因組異常通常在FF環境中作為單細胞培養的hESC和hiPSC系中檢測到(Laurent等人,2011;Taapken等人,2011)。所有分析的hiPSC系的核型分析證實在FMM培養中的基因組穩定性(圖4A)。另外,在FMM中維持延長時期(25-30代)的單細胞和FF培養的hiPSC克隆繼續維持其未分化概況和基因組穩定性,而無需培養物清除或選擇(圖4B)。
在FMM中維持的附加體衍生的hiPSC克隆也容易產生所有三個體細胞譜系,經由胚狀體的體外分化,以及通過畸胎瘤形成的體內分化(圖4C-E)。
這些數據證實FRM和FMM多階段培養基平臺使無轉基因hiPSC克隆能夠以FF和單細胞酶促傳代形式容易生成且擴增,同時維持多能和基因組穩定的細胞的同質群。
實例4
多階段培養基平臺使使用最低限度基因的附加體重編程成為可能
概述
具有癌基因例如KLF4、MYC和LIN28的減少依賴性或在重編程過程中需要敲減P53的有效的無足跡表達系統具有用於多能幹細胞治療的極大價值(Lee等人,2013;Okita等人,2011;Yu等人,2009)。因為多階段培養基平臺證實使用OSNKLMT重編程因子用於生成和擴增無轉基因hiPSC細胞的極度有效和穩健的平臺,平臺的穩健性針對重編程因子的需求進行測量。
結果
在改變基因表達組合和劑量的嘗試中,構建包含最低限度基因集合的幾種附加體表達盒,包括OCT4/NANOG/SOX2(ONS)、OCT4/SOX2(OS)或OCT4/OCT4(2xO)(圖5A)。用OCT4、NANOG、SOX2和SV40LT的組合轉染成纖維細胞細胞系,並且使用FRM/FMM平臺進行培養。在重編程第13天時,SV40LT單獨不產生任何真正的SSEA4/TRA1-81陽性細胞,但改善轉染後的細胞存活(圖5B和11A)。各種重編程因子組合導致有效重編程,如通過SSEA4/TRA1-81陽性群在重編程過程中早期的出現證實的(到第13天時,關於OCT4/SOX2/SV40LT的>0.5%和關於OCT4/NANOG/SOX2/SV40LT的>3.5%;圖5B)。重編程另外幾天的培養物顯著增加SSEA4/TRA1-81陽性群(到第16天時,關於OCT4/SOX2/SV40LT的>4.0%;圖11B)。令人驚訝的是,觀察到的重編程細胞的百分比與含有癌基因KLF4和MYC的慢病毒和附加體誘導系統可比較(圖2B、5B和11B)。
在流式細胞術分選和選擇個別hiPSC克隆之前,幾種重編程因子組合推進且轉變為FMM培養基。類似於OSNKLMT附加體重編程,克隆hiPSC系容易來源於含有最低限度基因OCT4、SOX2和SV40LT的組合(2xO+OS+T)以及其他組合(圖5C)。到第5-7代時已喪失轉基因和附加型載體標記物的hiPSC克隆推進用於進一步分析(圖5D)。所選擇的克隆作為單細胞在FF環境中連續傳代,且維持基因組穩定的未分化細胞的同質群,且展示有效分化成三個體細胞譜系的能力(圖5E-J)。
總之,這些數據指示hiPSC通過在FRM/FMM、基於流式細胞術的重編程平臺中瞬時表達最低限度的重編程基因而容易地生成。
實例5
FMM平臺支持基態多能性
概述
為了進一步評估FMM平臺,通過現有方法重編程的體細胞的基因表達特徵與使用本文考慮的FMM平臺重編程的體細胞的基因表達特徵相比較。評估小分子和常規維持的hiPSC培養之間的基因表達差異(Hanna等人,2010b;Saha和Jaenisch,2009)。
結果
在一組實驗中,評估小分子介導的培養和常規培養之間的基因表達模式。慢病毒誘導的hiPSC克隆FTi111在小分子培養中生成且維持,且顯示為多能的。克隆直接解凍到各種培養環境內,包括:i)具有飼養細胞的常規培養基,和ii)具有飼養細胞或在FF表面上的含有小分子抑制劑的培養基(圖12A和12B)。在常規培養中的hiPSC集落僅在Thiazovivin,ROCK抑制劑的存在下回收,並且隨後將回收的細胞轉換為塊培養(圖12B和12C)。每組培養條件證實獨特的集落形態(圖12D)和多能標記物的基因表達的不同模式(圖12E)。在飼養細胞上的常規維持的培養比其在小分子中維持的配對培養更緊密地類似在常規培養中維持的hESC對照H1和HUES9(圖12E)。這些數據顯示不同的基因表達模式存在於小分子介導的培養和常規培養之間。
基因表達中的差異還在使用慢病毒誘導和常規ESC/飼養者培養衍生的hiPSC以及附加體衍生的系之間進行評估,並且進一步在FRM/FMM平臺中維持的用重編程因子的不同組合衍生的附加體系之間進行評估。高含量qRT-PCR分析用於定量與多能性和分化相關的基因表達。調查的大多數多能性基因展示在FMM培養或含有飼養細胞的常規培養中維持的hiPSC之間可比較的表達模式(圖6A)。然而,在與分化相關的基因的評價上觀察到細胞系之間的差異(圖6B)。當與在常規培養基中和在飼養細胞上維持的hiPSC和H1hESC兩者相比較時,FMM維持的hiPSC展示與三個體細胞譜系相關的大多數基因的更低表達。通過附加型基因集合OSNKLMT誘導的系亞集看起來顯示外胚層譜系OTX2和TUJ1的表達,而這種表達在不使用Lin28、KLF4和c-MYC的附加體衍生的hiPSC中可忽略不計(圖6B)。令人驚訝的是,測試的所有分化基因的表達在來源於附加體最低限度基因集合且在FMM中維持的所有hiPSC中被完全壓制(圖6B)。總之,這些數據指出FRM/FMM平臺可使用很少的基於附加體的重編程因子使細胞穩健地重編程,並且可使hiPSC維持在穩定多能基態。
對於來源於下述方法的hiPSC測定總體基因表達模式:i)在FMM中維持的附加體誘導,ii)在FMM中維持但對於三次傳代轉變為常規培養基的附加體誘導,iii)在SMC4中維持的慢病毒誘導;和iv)在常規培養中維持的慢病毒誘導(圖13A、13B)。在評估基因表達譜之前,所有系均測定為多能的、基因組穩定的;並且能夠分化成所有三個體細胞譜系。在小分子培養和常規培養之間差異表達的基因的簇分析揭示hiPSC系基於目前的培養條件而不是最初的衍生方法和培養成組(圖13B)。展示2.5倍表達差異的300種基因的基因本體分類將分化和發育鑑定為在常規培養組中高度富集的主要類別,而在小分子培養組中上調的基因與細胞增殖和性發育調節最相關(圖13C和13D)。
重複基因表達分析,並且直接比較FMM、常規或轉變培養系統(FMM,Conv,FMM→Conv,Conv→FMM;圖13A)。簇分析產生與生成方法或先前培養系統無關,基於目前培養系統分開的兩組(圖7A)。例如,在轉變為常規培養之前,hiPSC克隆FTC016-c28在FRM/FMM平臺下生成且維持。這個克隆與在常規培養中專一地維持的培養物而不是與其在FMM中維持的親本系成組;可比較結果用系例如OKS慢病毒誘導的hiPSC克隆可見,所述hiPSC克隆在常規培養中生成且在常規集合內成組直至轉變為FMM,在其上它在FMM簇內成組(圖7A)。基因本體使常規培養分類以富集與分化和發育相關的基因(即p值=2.2E-10,模式指定過程;圖7B和13E)。總之,這些數據顯示與分化傾向相關的基因在FMM培養中顯著減少,並且hiPSC可適合FMM培養平臺,以減少自發分化潛能。
匯集代表人多能幹細胞的基態和亞穩態的基因列表(De Los Angeles等人,2012;Han等人,2011;Hanna等人,2010a;Hirata等人,2012;Nichols和Smith,2012;Valamehr等人,2012;Zhou等人,2010)(圖7C)。對於在FMM或常規培養中的hiPSC系執行基於這些基因列表的基因聚簇(圖7D)。類似於總體基因表達比較,集中的基因聚簇顯示基於其目前培養條件的細胞系分開,具有看起來可相互轉換的概況。例如,從FMM轉變為常規培養的hiPSC克隆FTC016-c28與H1hESC而不是與其在FMM中維持的親本hiPSC系成組(圖7D)。類似地,來源於在常規培養中維持的成纖維細胞系的慢病毒hiPSC克隆與HUES9hESC和在常規培養中的其他hiPSC克隆成組;然而,當轉變為FMM時,它與來源於臍帶血的附加體hiPSC以及其他FMM培養的系成組(圖7D)。代表兩個簇內的基態和亞穩態的基因分布通過就小分子(SMC4/FMM)相對於常規培養而言標繪關於每個探針組的平均強度進行測定(圖7E)。令人驚訝的是,與基態相關的大多數基因顯示在小分子培養簇中升高的表達,並且在常規培養簇中檢測到與亞穩態相關的基因的增加表達(圖7E)。
在常規培養中培養且維持的hiPSC的X失活狀態與其適應小分子培養且維持10代的配對物相比較(圖7F)。當與常規培養相比較時,在小分子培養中維持的hiPSC顯示X染色體基因表達的增加,其提示沉默的X染色體的復活(圖7F)。值得注意的例外是X失活特異性轉錄物(XIST),其在轉變為小分子培養中是下調的(圖7F)。X活化的進一步證據通過相對於其適應常規培養基的配對培養,在FMM中培養的hiPSC中的H3K27me3的差異染色提供(圖7G)。在FMM中的大多數hiPSC缺乏H3K27me3染色;然而,在常規培養中的大多數hiPSC展示H3K27me3核灶,具有核大小減少的外觀,這提示X失活(與常規培養中的>90%H3K27me3染色相比較,在FMM中的<10%H3K27me3染色;圖7G)。
實例6
小分子培養中的hiPSC維持
一旦培養物的匯合達到75-90%,衍生的hiPSC(用重編程因子的各種組合包括OCT4/NANOG/SOX2、OCT4/ECAT1/UTF1或OCT4/ECAT1/UTF1/ESRRB/NANOG誘導的成纖維細胞或血細胞)就作為單細胞照常規傳代。對於單細胞解離,hiPSC用磷酸鹽緩衝鹽水(PBS)(Mediatech)洗滌一次,且在37℃下用Accutase(Millipore)處理3至5分鐘,隨後為吹吸以確保單細胞解離。單細胞懸浮液隨後以等體積與常規培養基混合,以225g離心4分鐘,重懸浮於命運維持培養基(FMM)中,且在hESC合格Matrigel(Corning)塗布表面上鋪平板。根據製造商的說明書,製備Matrigel且用於塗布表面。傳代通常為1:3-1:6,組織培養板先前在37℃下用Matrigel塗布1-4小時,並且每兩到三天用FMM供養。細胞培養在設為37℃和5%CO2下的加溼溫箱中維持。常規培養基由DMEM/F12(Mediatech)、20%敲除血清替代物(Life Technologies)、1x Glutagro(Mediatech)、1x非必需胺基酸(NEAA)(Mediatech)、1x Pen/Strep(Mediatech)和100μMβ-巰基乙醇組成。FMM由常規培養基組成,所述常規培養基補充有5μM Thiazovivin(內部合成)、0.4μM PD0325901(Biovision)、1μM CHIR99021(Biovision)、100ng/mLbFGF(Life Technologies)和10ng/mL hLIF(Millipore)。關於在FMM中擴增的培養物的流動分析和形態呈現於圖18A-D和19A-B中。在FMM中擴增的細胞還證實經過多次傳代的正常核型,如圖20A-D中所示。
實例7
在小分子培養中使用最低限度基因的重編程
為了起始重編程,重編程因子的後生表達通過使用NIL的慢病毒轉導、常規整合慢病毒或用附加型載體的電穿孔進行誘導。如圖14A-B中所示,慢病毒表達系統由幾個特點組成,所述特點包括EF1α啟動子、特異性基因組合(表3)和在3'末端處的LOXP位點,以允許CRE介導的整合轉基因的切除。在CRE切除後,衍生的hiPSC基因組不再含有轉基因且基本上是無足跡的。如圖14C-F中所示,附加型構建體具有獨特特點,包括EF1α啟動子和獨特的重編程因子。在轉染後,附加型構建體位於核內,並且以不整合到基因組內的反式介導方式起作用。
對於慢病毒感染,根據製造商的說明書,起始人成纖維細胞以7x104-1x105個細胞/用Matrigel(Corning)塗布的6孔板的孔進行種植。來自293T細胞的新鮮慢病毒上清液以1∶2稀釋度(一份慢病毒上清液:一份成纖維細胞培養基)加入起始細胞中。NIL病毒上清液以1x濃度使用且不稀釋。如果使用先前冷凍的病毒,則它不稀釋且以1x濃度使用。各種因子的病毒上清液組合(表3)直到總共2mL培養基/6孔。這補充有5μg/mL聚凝胺(Millipore)和10mM Hepes(Meditech),隨後為自旋感染。六孔板用石蠟膜密封且在32℃下以600g離心90分鐘。板隨後轉移至37℃和5%CO2溫箱12-16小時。在與慢病毒一起溫育後,細胞用PBS進行洗滌,並且培養基轉變為含有一份命運重編程培養基(FRM)和一份成纖維細胞培養基的50/50培養基。培養基在感染後4至6天之間完全轉變為FRM。FRM由補充有下述的常規培養基(上文描述的)組成:5μM Thiazovivin(內部合成)、0.4μM PD0325901(Biovision)、1μM CHIR99021(Biovision)、2μM SB431542(Biovision)、100ng/mL bFGF(Life Technologies)、10ng/mL hLIF(Millipore)、1x N2補充物(Life Technologies)和1x B27補充物(Life Technologies)。一旦孔變得匯合,細胞就在先前用Matrigel塗布的10cm皿上進行傳代。傳代由在Matrigel塗布的表面上用Accutase(Millipore)解離組成(如上所述)。在第14和18天之間,或當iPSC集落變得存在時,培養基從FRM轉變為FMM。單細胞解離的細胞擴增到具有FMM的Matrigel塗布的板上,並且維持直至流式細胞術分選時。關於hiPSC表型表達的結果呈現於表3、圖15A-C(在8-15天時)和圖16A-D、圖17A-B(在第4周時Oct-4/Ecat1/UTF1/Esrrb/Nanog)。
表3:重編程因子組合和多能表型表達
對於附加型載體重編程,使用NEON轉染系統(Life Technologies)進行使用圖13中所示的質粒的成纖維細胞或臍帶血細胞轉染。如通過產品手冊描述的,大致地,使用在適當緩衝液中1650v/10ms/3次脈衝的設置,總共3μg含有重編程因子的附加型質粒與EBNA(以mRNA的形式或作為克隆質粒pCDNA中的盒)共轉染到5x105成纖維細胞或2.5x105臍帶血細胞內。將經轉染的細胞直接種植到6孔板的孔上,所述6孔板用含有Matrigel的成纖維細胞培養基或臍帶血培養基(取決於細胞類型)進行塗布,所述培養基補充有4ng/mL bFGF和5μg/mL纖連蛋白(BD Biosciences),不含抗生素。臍帶血培養基由SFMII+CC110(Stem Cell Technologies)組成。轉染後二十四小時,將FRM以等體積加入培養中。對於成纖維細胞培養,轉染後四十八小時,將50μg/mL潮黴素(Corning)加入培養中。培養基在第5天時完全轉變為FRM,其中潮黴素在轉染後7天去除。轉染後14天,所有重編程培養轉變為FMM。對於臍帶血培養,感染後二十四小時,將FRM以等體積加入,並且每幾天連續加入,直至轉染後第14天,其中培養物被抽吸且替換為完全FMM。在兩種情況下,在轉染後5至7天左右可見貼壁圓形細胞的簇。一旦在FMM中,所有重編程培養均得到維持,並且在Matrigel塗布的表面上使用Accutase進行單細胞傳代(上文描述)。單細胞解離的細胞擴增到具有FMM的Matrigel塗布的板上,並且維持直至流式細胞術分選。
實例8
重編程因子及其化學計量學的影響
人成纖維細胞用含幾種重編程因子的慢病毒進行自旋感染。所有樣品均用OCT4、SOX2、NANOG和SV40LT進行感染,使用不含抗生素選擇因子的慢病毒質粒。細胞用含有嘌呤黴素選擇盒以及各種重編程因子的單一慢病毒質粒共感染。這些因子包括OCT4-P2A-SOX2、OCT4-P2A-NANOG-T2A-SOX2或OCT4-P2A-OCT4。感染後兩天,將在50/50培養基中的500ng/mL嘌呤黴素(Life Technologies)加入每個孔中。在第5天時,在嘌呤黴素選擇的三天後,培養基變成不含嘌呤黴素的FRM。在第14天時,培養基轉變為FMM。在第24和27天之間,對於SSEA4+/TRA181+群進行流動分析。觀察到增加的OCT4表達顯著改善重編程效率(圖23A)。
實例9
實驗程序
在常規培養系統中的hiPSC維持
常規培養的hiPSC維持在絲裂黴素C處理的MEF(Millipore)飼養細胞上,並且用常規培養基(在正文中被稱為常規培養基)進行培養,所述常規培養基含有DMEM/F12(Mediatech)、20%v/v敲除血清替代物(Life Technologies)、1%v/v非必需胺基酸(Mediatech)、2mM L-穀氨醯胺(Mediatech)、100μMβ-巰基乙醇(Life Technologies)和10ng/mL bFGF(Life Technologies)。在匯合時,常規培養的hiPSC使用1mg/mL膠原酶IV(Life Technologies)在37℃下酶促解離7分鐘,隨後機械解離成小片(稱為塊傳代),收集且每5–7天1:3–1:4稀釋傳代到新鮮種植的飼養細胞上,伴隨常規培養基的每天添加。在過度自發分化的情況下,手動挑選集落且使用胰島素注射器(Becton Dickinson)的尖端切割成小片,並且轉移至新鮮種植的飼養細胞。細胞培養維持在設為37℃和5%CO2下的加溼溫箱中。
體細胞的重編程
為了起始重編程,重編程因子的後生表達通過慢病毒轉導或附加型載體轉染進行誘導。慢病毒轉染遵循如先前描述的(Valamehr等人,2012)。簡言之,起始細胞以1x105個細胞/6孔板的孔鋪平板到Matrigel(BD Biosciences)塗布的表面上。除非指定,否則所有Matrigel塗層均由下述組成:將Matrigel溶液(重懸浮於25mL DMEM/F12中的1等分試樣的Matrigel)加入組織培養表面,並且允許在37℃下的2-4小時溫育。來自生成表達轉基因OCT4/SOX2/KLF4的慢病毒的293T細胞的上清液以1∶2稀釋度(一份慢病毒上清液:1份成纖維細胞培養基)加入起始細胞,補充有4μg/mL聚凝胺(Millipore),並且轉移至37℃和5%CO2共12-16小時。成纖維細胞培養基:DMEM(Mediatech)、10%FBS(Life Technologies)、1x glutamax(Life Technologies)、1x非必需胺基酸(Mediatech)。在與慢病毒一起溫育後,細胞用PBS洗滌三次且用成纖維細胞培養基供養。轉染後48小時,培養基轉變為含有一份FRM(或SMC4)和一份成纖維細胞培養基的50/50培養基。一旦培養物傳代到較大容器內,通常在轉染後第4至6天之間,培養基就完全轉變為FRM(或SMC4)。傳代由在Matrigel塗布的表面上用Accutase解離組成(如下所述)。培養維持在FRM(或SMC4)中直至下一次應用。
對於附加型載體重編程,使用NEON轉染系統(Life Technologies)進行使用基因集合OCT4/SOX2/NANOG/KLF4/LIN28/MYC/SV40LT(A14703,Life Technologies)的成纖維細胞或臍帶血細胞的轉染。如通過產品手冊描述的,大致地,使用在適當緩衝液中1650v/10ms/3次脈衝的設置,將4μg載體組轉染到5x105個成纖維細胞或2.5x105個臍帶血細胞內。將經轉染的細胞直接鋪平板到10cm皿(成纖維細胞)或6孔板(臍帶血)的孔內,所述10cm皿或6孔板用Matrigel進行塗布,且含有成纖維細胞培養基或臍帶血培養基(取決於細胞類型),所述培養基補充有10ng/mL bFGF和5μg/mL纖連蛋白(BD Biosciences)。臍帶血培養基:SFMII+CC110(Stem Cell Technologies)。轉染後二十四小時,將FRM以等體積加入培養中。對於成纖維細胞培養,轉染後四十八小時,將50μg/mL潮黴素(Mediatech)加入培養中。培養基在第5天時轉變為完全FRM,其中潮黴素在轉染後第7天時去除。在轉染後第14天時,所有重編程培養轉變為FMM。對於臍帶血培養,轉染後二十四小時,將FRM以等體積加入,並且每幾天連續加入,直至轉染後第14天,其中培養物被抽吸且替換為完全FMM。在兩種情況下,在轉染後5至7天左右可見貼壁圓形細胞的簇。一旦在FMM中,所有重編程培養均得到維持,並且使用Accutase進行單細胞傳代。單細胞解離的細胞擴增到具有FMM的Matrigel塗布的板上,並且維持直至流式細胞術分選。在玻連蛋白(Life Technologies)表面塗布研究中,所有方面均保持相同,除了將Matrigel替換為玻連蛋白之外。對於減少因子的附加體重編程,pCEP4(Life Technologies)載體主鏈構建為含有在EF1α啟動子調節下的OCT4-P2A-OCT4、OCT4-P2A-SOX2或OCT4-P2A-NANOG-T2A-SOX2。減少因子的附加型載體的轉染遵循與上文所述相同的方案,除了少數修飾之外。EBNA與EBNA mRNA(20μg)或載體盒(2μg)共轉染(Howden等人,2006)。潮黴素選擇維持10天,並且FMM在第16天時引入。
慢病毒的生成
293T(ATCC)維持在不含抗生素的成纖維細胞培養基中,且允許達到超過80%匯合。成纖維細胞培養基由DMEM(Mediatech)、10%胎牛血清(FBS)(Life Technologies)、1x Glutagro(Mediatech)和1x NEAA(Mediatech)組成。細胞通過首先用PBS洗滌隨後在37℃下與0.05%胰蛋白酶(Mediatech)一起的4分鐘溫育進行傳代。解離的細胞重懸浮於成纖維細胞培養基中,以225g離心4分鐘,且種植到所需板上。為了生成整合慢病毒,對於每種病毒製備物,293T在第1天時以3.5x106個細胞/10cm皿進行傳代。在第2天時,轉染前1小時,培養基變成10mL新鮮成纖維細胞培養基。DNA使用CalPhos Kit(Clontech)進行轉染。將下述組合用於轉染:5μg含有目的基因的慢病毒克隆質粒、3.2μg包裝質粒pPAX、630ng包裝質粒pMDG、87μL鈣溶液和水直到700μL。加入700μL HBS溶液,同時使用1mL血清學吸管產生氣泡。這在室溫下溫育15分鐘,且隨後逐滴加入293T的10cm板。在第3天時,取出病毒上清液,棄去,並且將15mL新鮮成纖維細胞培養基加入板中。在第4天時,轉染後48小時,收集病毒上清液且貯存於4℃下。將15mL成纖維細胞培養基加入板中。在第5天時,轉染後72小時,收集病毒上清液且加入第4天上清液中。這個病毒合併物(pool)使用0.45μm濾器進行過濾,並且使用Lenti-X GoStix(Clontech)檢查滴度。病毒用於感染或以等分試樣在-80℃下冷凍。為了生成非整合慢病毒(NIL)(Invivogen),對於每種病毒製備物,使用T75燒瓶,遵循根據製造商說明書的方案。轉染後48、72和96小時收集病毒上清液,合併,過濾且如上所述滴定。NIL病毒用於感染或以等分試樣在-80℃下冷凍。
在小分子培養中的hiPSC維持
一旦培養物的匯合達到75-90%,衍生的hiPSC就照常規作為單細胞進行傳代。注意到過度匯合可導致分化。對於單細胞解離,hiPSC用磷酸鹽緩衝鹽水(PBS)(Mediatech)洗滌一次,且在37℃下用Accutase處理3至5分鐘,隨後為吹吸以確保單細胞解離。單細胞懸浮液隨後以等體積與常規培養基混合,以225g離心4分鐘,重懸浮於FMM中,且在Matrigel塗布的表面上鋪平板。傳代通常為1:4–1:8,轉移先前在37℃下用Matrigel塗布2-4小時的組織培養板,並且每隔一天用FMM供養。細胞培養在設為37℃和5%CO2下的加溼溫箱中維持。關於FMM和FRM的培養基配方在表1中描述。先前討論了SMC4培養(Valamehr等人,2012)。簡言之,將小分子0.4mM PD0325901(Biovision)、1mM CHIR99021(Biovision)、5mM Thiazovivin和2mM SB431542(Biovision)加入常規培養基中,並且根據方案進行傳代。
流式細胞術分析和分選
單細胞解離的(上文描述)重編程庫重懸浮於冷卻的染色緩衝液中,所述染色緩衝液含有Hanks'平衡鹽溶液(MediaTech)、4%胎牛血清(Invitrogen)、1x青黴素/鏈黴素(Mediatech)和10mM Hepes(Mediatech)。將綴合的一抗包括SSEA4-FITC、TRA1-81-Alexa Fluor-647和CD30-PE(BD Biosciences)加入細胞溶液中,且在冰上溫育15分鐘。所有抗體均在100μL染色緩衝液以7-10μL/百萬個細胞使用。溶液在染色緩衝液中洗滌一次,以225g向下旋轉4分鐘,且重懸浮於含有10μM Thiazovivin的染色緩衝液中,且在冰上維持用於流式細胞術分選。使用同上描述的門控策略,在FACS Aria II(BD Biosciences)上執行流式細胞術分選。分選的細胞使用100μM噴嘴以3和9個事件/孔的濃度直接噴射到96孔板內。分選3個細胞/孔是優選濃度,因為注意到分選的事件不一定與分選後每個孔中可見的實際細胞數目關聯,並且3個細胞/孔給予含有個別集落的優選孔數目。每個孔預填充有補充了5μg/mL纖連蛋白和1x青黴素/鏈黴素(Mediatech)的200μL FMM,並且先前用5x Matrigel塗布過夜。5x Matrigel預塗布包括將一等分試樣的Matrigel加入5mL DMEM/F12內,隨後在4℃下溫育過夜,以允許適當重懸浮且最後以50μL/孔加入96孔板,隨後為在37℃下的過夜溫育。在將培養基加入每個孔之前立即抽吸5x Matrigel。在分選完成後,96孔板在溫育前以225g離心1-2分鐘。板不受打擾七天。在第七天時,從每個孔中取出150μL培養基且替換為100μL FMM。孔在分選後第10天時用另外100μL FMM再供養。集落形成早在第2天時檢測到,並且大多數集落在分選後第7-10天之間進行擴增。在第一代中,孔用PBS洗滌且在37℃下用30μL Accutase解離大約10分鐘。延長的Accutase處理的需要反映在培養中靜置延長持續時間的集落的緊密性。在細胞可見解離後,將200μL FMM加入每個孔中,並且吹吸幾次以破壞集落。解離的集落轉移至先前用5x Matrigel塗布的96孔板的另一個孔,並且隨後在溫育前以225g離心2分鐘。進行這種1:1傳代以使早期集落鋪展開。後續傳代照常規完成:用Accutase處理3-5分鐘,並且1:4擴增到先前用在FMM中的1x Matrigel塗布的更大的孔內。流式細胞術分析在Guava EasyCyte 8HT(Millipore)上執行,並且使用FCS Express 4(De Novo Software)進行分析。
實時RT-PCR和Fluidigm分析
使用Pico Pure RNA Isolation Kit(Life Technologies)分離總RNA。使用iScript cDNA Synthesis Kit(Bio-Rad),由100ng分離的總RNA逆轉錄互補DNA(cDNA)。使用TaqMan PreAmp Master Mix Kit(Life Technologies)和0.2x濃度的合併TaqMan測定,cDNA隨後用於22種特異性靶基因和兩種參考對照基因的預擴增。使用14個擴增循環,使用在製造商的方案中所述的標準循環條件,執行來自cDNA的特異性靶擴增(STA)。預擴增的cDNA反應(n=48)1:5稀釋(在無菌水中),並且用作實時定量PCR反應的模板。使用NanoFlex IFC Controller MX(Fluidigm)裝載48.48Dynamic陣列(Fluidigm),其中TaqMan測定一式兩份裝載,並且使用BioMark Real-Time PCR System(Fluidigm)執行實時反應。使用BioMark Real-Time PCR Analysis軟體(Fluidigm)分析結果。從計算中排除具有超過32的循環閾值(Ct)的樣品。平均Ct由測定重複進行計算,並且使用兩種參考基因(GAPDH和HPRT1)的平均值針對六種對照MEF細胞系(在MEF上的OSK hiPSC和H1ESC)的中值計算delta-delta Ct(ΔΔCt)。相對基因表達(RQ)結果以熱圖形式在Excel(Microsoft)中展示。
表4.FAM標記的TaqMan探針
測試轉基因的存在
使用DNA Mini Kit和蛋白酶K消化(Qiagen)分離基因組DNA。使用Taq PCR Master Mix Kit(Qiagen),使用轉基因特異性引物組(下表5)擴增100ng基因組DNA(Yu等人,2007)。PCR反應如下運行35個循環:94℃30秒(變性)、60-64℃30秒(退火)和72℃1分鐘(延伸)。來自使用慢病毒方法生成的成纖維細胞和hiPSC的基因組DNA用作陰性對照。附加型構建體的DNA用作陽性對照。
表5.轉基因特異性引物組
免疫細胞化學分析
細胞使用4%v/v多聚甲醛(Alfa Aesar)進行固定,用含有0.2%v/v Tween的PBS(PBST)(Fisher Scientific)洗滌三次,並且在25℃下使用在PBS中的0.15%v/v TritonX-100(Sigma-Aldrich)滲透化處理1小時。在滲透化處理後,細胞在25℃下用在PBST中的1%v/v BSA(Invitrogen)(PBSTB)(Fisher Scientific)封閉30分鐘。在PBSTB的輕柔去除後,細胞與在PBSTB中的一抗一起在4℃下溫育過夜。在該研究中使用的一抗包括OCT4(Santa Cruz)、NANOG(Santa Cruz)、TRA160(Millipore)、TRA1-81(Millipore)、SSEA4(Millipore)、β-III微管蛋白(TUJ1、R&D Systems)、α-平滑肌肌動蛋白(Sigma)、FoxA2(R&D Systems)、Sox17(R&D Systems)、NESTIN(Abcam)和α-1-胎蛋白(Dako)。在過夜溫育後,細胞用PBST洗滌三次,並且在37℃下用在PBSTB中1:250稀釋的二抗(Alexa Fluor 488或555;Invitrogen)染色1小時。細胞在PBS中洗滌三次,並且用Hoechst染料(Invitrogen)進行染色。對於H3K27me3染色分析,hiPSC在蓋玻片上生長72至96小時,並且在25℃下用在PBS中的4%多聚甲醛(Electron Microscopy Science,EMS)固定15分鐘。細胞滲透化處理在25℃下用在PBS中的0.1%Triton X-100執行1小時,並且隨後細胞在25℃下與封閉溶液(在PBS中的1%BSA)一起溫育30分鐘。在封閉後,蓋玻片在4℃下與抗三甲基組蛋白H3(Lys27)抗體(Millipore 07-449,H3K27me3)的1:1600稀釋物一起在封閉溶液中溫育過夜。二抗是Alexa Fluor 555山羊抗兔IgG(Life Technologies,A21429)。核用DAPI進行復染,並且用Axio Observer Inverted Microscope(Carl Zeiss)觀察。用AxioVS40v4.8.1.0(Carl Zeiss Imaging Solutions Gmbh)捕獲圖像。
根據實例7重編程的細胞使用4%v/v多聚甲醛(Alfa Aesar)進行固定,用PBS(Mediatech)洗滌,並且在25℃下使用在PBS中的0.15%v/vTritonX-100(Sigma-Aldrich)滲透化處理1小時。在滲透化處理後,細胞在25℃下用在PBS中的1%v/v BSA(Sigma)(PBSB)封閉30分鐘。在PBSB的輕柔去除後,細胞與在PBSB中的一抗一起在4℃下溫育過夜。在該研究中使用的一抗包括OCT4(Santa Cruz)和TRA181(Millipore)。在過夜溫育後,細胞用PBS洗滌三次,並且在37℃下用在PBSB中1:250稀釋的二抗(Alexa Fluor 488或555;Life Technologies)染色1小時。細胞在PBS中洗滌三次,並且用Hoechst染料(Invitrogen)進行染色。染色的細胞用Axio Observer Inverted Microscope(Carl Zeiss)觀察。用AxioVS40v4.8.1.0(Carl Zeiss Imaging Solutions Gmbh)捕獲圖像。
分化分析(EB和定向)
hiPSC在分化培養基中分化為EB,所述分化培養基含有DMEM/F12(Mediatech)、20%胎牛血清(Invitrogen)、1%非必需胺基酸(Mediatech)、2mML-穀氨醯胺(Mediatech)和100μMβ-巰基乙醇。簡言之,對於EB形成,將hiPSC種植到FMM中,並且第二天轉變為常規,以使細胞引發。在常規培養基中3至4天後,培養物是用Accutase(Millipore)解離的單細胞,並且在包括10μM Y27632的分化培養基中重懸浮至100,000個細胞/mL的最終濃度。注意到ROCK抑制劑Y27632代替Thiazovivin用於EB形成。細胞以100μL/孔種植到V形底96孔非組織培養板(Nunc)中,並且以950g離心5分鐘。第二天,使用P1000以在分化培養基中的大約30–40個EB/孔,將緊密的「球樣塊」轉移至超低結合6孔板(Corning)。在7天後,EB以1:1轉移至Matrigel塗布的6孔板,並且每三天用分化培養基供養。在培養3周後,將細胞固定且染色。對於定向單層分化,將hiPSC種植到Matrigel塗布的孔中的FMM中,以在第二天遞送50%和90%匯合。兩個密度均誘導至分化。對於神經誘導,將FMM培養基替換為補充有10μM SB431542和100nM LDN-193189(兩種SMAD抑制劑,Biovision)的hESC培養基。在2天後,除雙重SMAD抑制劑之外,分化培養基還補充有3μM CHIR99021(Biovision)。兩天後使細胞固定且就巢蛋白(Abcam)進行染色。對於中胚層分化,將培養基替換為補充有1x B27培養基添加劑(Life Technologies)、3μM CHIR99021、4ng/ml bFGF和10ng/ml BMP4的RPMI(Mediatech)。每隔一天更換培養基,並且在第4天時使細胞固定,並且就αSMA(Sigma)進行染色。使用Human Pluripotent Stem Cell Functional Identification Kit(R&D Systems)執行內胚層分化。hiPSC與內胚層分化培養基一起溫育3天,固定且就SOX17(R&D Systems)進行染色。
基因表達分析
使用製造商推薦的方案,使用PicoPure RNA Isolation試劑盒(Life Technologies)提取RNA。使用Nanodrop 2000分光光度計(Thermo Scientific)定量總RNA。簡言之,使用用於MessageAmp II aRNA Amplification Kit(Applied Biosystems/Ambion,Austin,TX)的標準方案,利用任選的第二輪擴增,由大致100ng總RNA製備生物素化的aRNA,並且隨後使用標準方案,使用MessageAmp II Biotin Enhanced Kit(Applied Biosystems/Ambion,Austin,TX),轉錄成生物素標記的aRNA。生物素標記的aRNA根據Affymetrix建議進行純化且片段化。20μg片段化的aRNA在45℃下用於與Human Genome U133-plus-2.0晶片(Affymetrix Inc.Santa Clara,CA)雜交16小時。將陣列洗滌且在Affymetrix Fluidics Station 450中染色,並且使用Affymetrix GeneChip Scanner 3000 7G進行掃描。原始表達數據文件在Gene Expression Omnibus(GSE50868)上可獲得。使用Affymetrix Expression Console軟體,使用預設分析設置分析圖像數據。陣列通過對數標度的穩健的多陣列分析(RMA,Affymetrix)進行標準化,並且在Spotfire for Genomics 4.5(Tibco Spotfire,PaloAlto,CA)中顯現。差異表達的探針的生物途徑富集分析針對Gene Ontology(GO)資料庫(對GO生物過程的單獨富集和p值<2.5倍)或限定基態或亞穩態的靶向基因列表的存在加以選擇。對於X染色體基因表達比較,通過獲得2^[RMA log2強度],將RMA標準化的Affymetrix基因晶片強度轉換為線性表達值。線性表達比作為幼稚表達集合除以引發的表達集合進行計算。關於對X染色體作圖的所有探針組的表達比在Spotfire 4.5中顯現,其中大於或小於2倍富集比的探針組是突出顯示的。
核型分析
通過WiCell Research Institute(Madison,WI)對二十至四十個G帶中期細胞執行細胞遺傳學分析。
畸胎瘤形成
以0.5和3百萬個細胞/200μL溶液(100μL FMM和100μL Matrigel)濃度的單細胞解離的hiPSC皮下注射到NOD/SCID/γ裸鼠內。在5-6周(3百萬個細胞注射)和7-8周(0.5百萬個細胞注射)後,畸胎瘤在PBS中收穫,在室溫下在4%多聚甲醛中固定過夜,並且其後在室溫下在70%乙醇中維持用於處理。將樣品提交給UCSD Histology Core Facility用於切片以及蘇木精與伊紅染色。使用配備Nikon DS-Fi1照相機的Nikon Eclipse TS100顯微鏡,將切片檢查,解釋且拍照。
統計分析
斯氏t檢驗用於關於標準差的統計評估。StepOne Software v2.2(Life Technologies)用於測定關於qRTPCR數據的RQ最小值和最大值(誤差條)。
一般而言,在下述權利要求中,所使用的術語不應解釋為使權利要求限制於說明書和權利要求中公開的具體實施例,而是應解釋為包括所有可能實施例連同此類權利要求賦予權力的等價物的完全範圍。相應地,權利要求不受公開內容限制。