一種永生化兔小腸上皮細胞系及其構建方法與流程
2023-09-17 00:19:30 2
本發明涉及分子生物學領域,具體地說,涉及一種永生化兔小腸上皮細胞系及其構建方法。
背景技術:
兔是一種重要的肉製品和皮毛來源,在畜牧生產、餐飲、公共健康等領域都有重要作用。目前,兔的重大疫病如兔瘟、兔球蟲、巴氏桿菌病等越來越受到重視。這類病原體多數都侵害消化道。然而,目前的研究中,研究這些疾病的科學工具嚴重缺乏,制約了相關領域的技術發展。目前兔來源的細胞系較少,因此獲得更多兔來源的細胞系將促進兔重大疫病防控的研究。
小腸來源的細胞系是研究各類腸侵襲性病原的可靠細胞學工具,而小腸隱窩來源的帶有lgr5的隱窩幹細胞更是研究小腸上皮發育分化的有利工具。因此亟需開發一種可獲得隱窩來源的永生化兔小腸上皮細胞系的方法,用以推進兔疫病防控和細胞學方面的研究。
技術實現要素:
為了解決現有技術中存在的問題,本發明的目的是提供一種永生化兔小腸上皮細胞系及其構建方法。
為了實現本發明目的,本發明的技術方案如下:
第一方面,本發明提供一種永生化兔小腸上皮細胞系的構建方法,採集新生兔的小腸,利用含有膠原酶和中性蛋白酶的酶消化液進行消化,消化後使用重懸離心和相差貼壁的方法對細胞進行提純,獲得原代隱窩細胞,進行培養與傳代,對獲得的細胞轉染sv40大t抗原質粒,再連續細胞傳代篩選得到永生化兔小腸上皮細胞系。
作為優選,所述酶消化液為含有0.1~0.2mg/ml膠原酶和0.4~0.6mg/ml中性蛋白酶的dmem培養液。例如,所述酶消化液為含有0.2mg/ml膠原酶和0.5mg/ml中性蛋白酶的dmem培養液。
進一步地,所述sv40大t抗原質粒包括雞球蛋白/兔球蛋白混合啟動子,sv40大t抗原基因和neo藥篩基因。所述sv40大t抗原質粒的構建方法為:將sv40大t抗原基因(geneid29031019)和neo藥篩基因(ng-047417.1)按順序插入到pcaggs載體內。
需要說明的是,目的基因插入質粒的順序不限於上述順序。所述pcaggs載體為市售可得商品,例如可購自belgianco-ordinatedcollectionsofmicro-organisms(bccm),產品代碼為lmbp2453(http://bccm.belspo.be/catalogues/lmbp-plasmids-plasmid-details?nm=pcaggs)。
進一步地,在所述培養與傳代的過程中,所述原代隱窩細胞進行初代培養時的培養基為添加10%fbs,20μg/l表皮生長因子,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑;gibco,貨號1805428)的dmem培養基。
更為具體地,本發明所述方法包括如下步驟:
(1)細胞分離:在無菌條件下取新生兔的小腸,切碎腸組織,dmem培養基衝洗腸組織2~3遍,使用上述含有膠原酶和中性蛋白酶的酶消化液在37℃下對腸組織進行消化,消化後使用重懸離心和相差貼壁方法對細胞進行提純,獲得原代隱窩細胞團;
(2)細胞培養:使用添加10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑)的dmem培養基,在包被有質量比0.5%明膠(sigma,貨號#slbn4365v)的培養板上對原代隱窩細胞進行初代培養;
(3)細胞傳代:當細胞團塊貼壁後生長至90%鋪滿狀態時進行細胞傳代,使用pbs洗滌去除培養基後,加入含質量比(m/m)0.02%edta的0.25%胰酶消化液進行消化,消化後將細胞重懸傳代到新的細胞瓶內,添加適量新細胞培養液;所述含質量比(m/m)0.02%edta的0.25%胰酶消化液為市售產品(macgene,中國;貨號g270220);
(4)細胞轉染:當細胞密度到達80%,鋪板時間36小時內,使用脂質體(lipofectamineltx,invitrogen公司,貨號1657815)轉染sv40大t抗原質粒,再連續細胞傳代,最終篩選得到永生化兔小腸上皮細胞系。
其中,步驟(3)中第1代到第5代使用的細胞培養液為10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑)的dmem培養基;第五代後使用的細胞培養液為7.5%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑)的dmem培養基。
第二方面,本發明還提供了利用本發明所述方法建立得到一種永生化兔小腸上皮細胞系。
該永生化兔小腸上皮細胞系可同時表達腸上皮表面標誌分子和隱窩基層幹細胞的表面分子。所述腸上皮表面標誌分子包括但不限於角蛋白8,e-降鈣素和ki67;所述隱窩基層幹細胞的表面分子包括但不限於lgr5,sox9。
第三方面,本發明還提供了所述永生化兔小腸上皮細胞系在建立兔小腸上皮體外模型中的應用,以及用於營養吸收,細胞通路,病原體研究等方向的研究。
本發明涉及到的原料或試劑,如無特殊說明均為普通市售產品,涉及到的操作如無特殊說明均為本領域常規操作。
在符合本領域常識的基礎上,上述各優選條件,可以相互組合,得到具體實施方式。
本發明的有益效果在於:
本發明提供了一種兔小腸隱窩來源的細胞系,該細胞系表達多種腸上皮細胞的表面標誌性分子,可以用於細胞生物學,病原與宿主相互作用的體外研究。該細胞係為促進兔健康養殖及其疫病防控提供了很好的細胞學工具。同時也為體外藥物評估,生物製藥等諸多行業提供了一種潛在的工具。
附圖說明
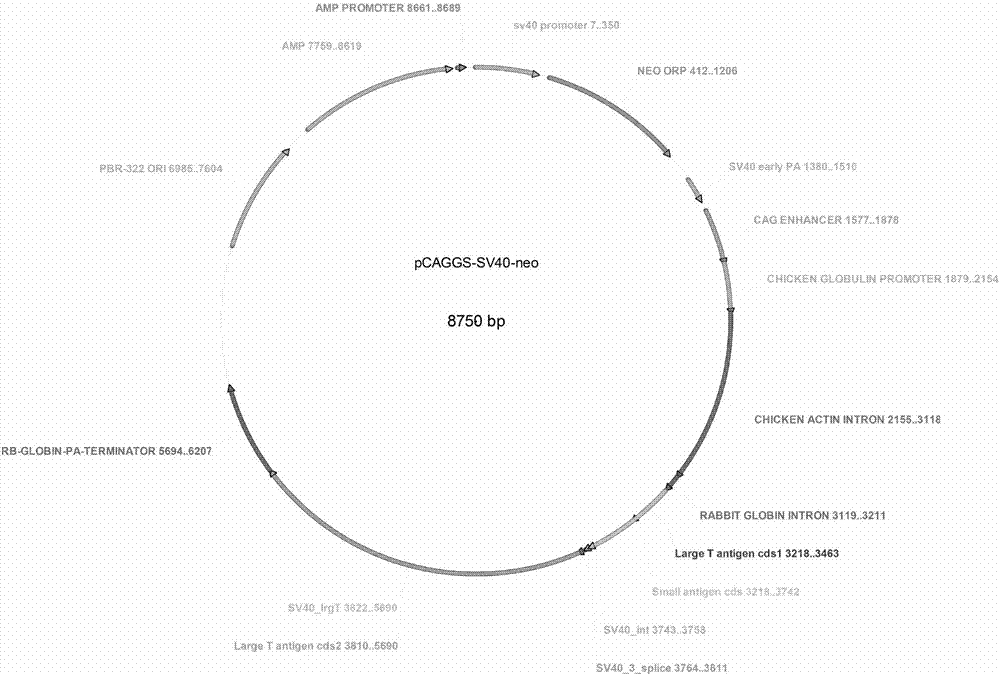
圖1為本發明所述sv40大t抗原質粒圖譜。
圖2為本發明所述永生化兔小腸上皮細胞形態;a:第3代原代兔小腸上皮細胞傳代後12h(標尺=100μm);b:第50代永生化兔小腸上皮細胞傳代後10h(標尺=100μm);c:第3代原代兔小腸上皮細胞鋪滿培養瓶後形態(標尺=100μm);d:鋪滿培養瓶的第50代永生化兔小腸上皮細胞(標尺=100μm)。
圖3為本發明第50代永生化兔小腸上皮與第3代兔小腸上皮生長曲線。
圖4為rt-pcr鑑定sv40大t抗原在兔小腸上皮細胞內的表達;泳道1:轉染24h後rt-pcr擴增出sv40大t抗原基因目的條帶;泳道2:轉染48h後rt-pcr擴增出sv40大t抗原基因的目的條帶;泳道3:第65代兔小腸上皮細胞系擴增出sv40大t抗原基因的目的條帶。
圖5為本發明所述永生化兔小腸上皮細胞培養兔大型艾美耳球蟲;a:培養24h,子孢子發育為子孢子形狀的裂殖子(標尺=50μm);b:培養48h,可見4核裂殖子(標尺=10μm);c:培養48h後,可見第一代裂殖體(標尺=10μm);d,e:培養72h後,可見發育即將成熟的第二代裂殖體(d標尺=50μm,e標尺=30μm);f:培養96h,成熟的二代裂殖體;g:培養96h後,釋放出的第二代裂殖子(標尺=20μm);h:培養120h,發育中的第三代滋養體(標尺=15μm)。
具體實施方式
下面將結合實施例對本發明的優選實施方式進行詳細說明。需要理解的是以下實施例的給出僅是為了起到說明的目的,並不是用於對本發明的範圍進行限制。本領域的技術人員在不背離本發明的宗旨和精神的情況下,可以對本發明進行各種修改和替換。
下述實施例中所使用的實驗方法如無特殊說明,均為常規方法。
材料:新生兔來源於紐西蘭白兔
儀器與試劑:二氧化碳培養箱(thermo,中國),倒置顯微鏡(olympus,日本),超淨工作檯(中國蘇淨集團泰安公司),高壓蒸汽滅菌鍋(sanyo,日本),細胞培養板與細胞培養瓶(costar,美國),dmem(macgene,中國,貨號g1004200),mem199(macgene,中國;貨號g1702020),帶有質量比(m/m)0.02%edta的0.25%的胰酶(macgene,中國;貨號g270220),胎牛血清(macgene,中國;貨號g20uno2),insulin-transferrin-selenium(gibco,美國;貨號1805428),冷水魚皮明膠(sigma,美國;貨號#slbn4365v),膠原酶(sigma,美國;產品編號c9722),中性蛋白酶(sigma,美國;產品編號d4818),lipofectamineltx(invitrogen,美國;貨號1657815),表皮生長因子(sigma,美國;產品編號e5036)。其他所用的材料試劑,若無特殊說明均可從商業渠道獲得。
實施例1
1、兔小腸上皮隱窩細胞團的分離與培養
在超淨臺內無菌取新生兔的小腸細胞,使用無菌剪刀縱向剪開小腸,再使用無菌手術刀片切碎腸組織為2mm2的組織碎片。dmem培養基衝洗腸道組織2~3遍。使用酶消化液對腸組織在37℃下進行消化20min,消化過程中需要不斷攪拌消化液。消化完成後,使用孔徑為0.5mm的銅濾網對消化產物進行過濾。過濾液500g離心5min,棄去上清液。使用質量比(m/m)2.5%的山梨醇溶液對沉澱物進行重懸,懸液300g離心2分鐘,收集上清於新的離心管內,再使用質量比(m/m)2.5%山梨醇重懸沉澱物,重複2~3次。收集的所有重懸液400g離心3min,棄去上清,並繼續加入質量比(m/m)2.5%山梨醇進行4~6次離心。最後可獲得的沉澱物為高純度的小腸上皮隱窩細胞團塊。將沉澱物加入培養基重懸後放於細胞培養板內培養1h後,然後水平十字搖動培養板後吸取培養液轉移隱窩團塊。這樣可以使成纖維細胞率先貼壁,進一步對隱窩細胞進行提純。
2、細胞培養
原代兔小腸上皮培養需要使用質量比(m/m)0.5%明膠包被的培養板。具體方法為,在細胞培養板或者細胞培養瓶內加入適量質量比(m/m)0.5%的明膠溶液,37℃細胞培養箱內孵育2h後放入4℃冰箱保存。使用前吸出明膠液體,使用pbs清洗一遍即可。新分離的隱窩細胞經過相差貼壁處理後,接種於培養板內既可以正常培養。前5代細胞貼壁性較差,所有使用的細胞培養板或者培養瓶必須經過質量比0.5%明膠包被。使用的培養基為加入了10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑)的dmem培養基。
3、細胞傳代
待兔小腸上皮細胞鋪滿90%細胞培養板或細胞培養瓶就可以進行細胞傳代。棄去瓶中陳舊的培養基,使用無菌pbs洗滌3次,25cm2的細胞培養瓶加入500μl含有質量比(m/m)0.02%edta的0.25%胰酶,十字搖晃保證鋪滿細胞瓶。37℃溫箱孵育3min。取出細胞培養瓶,拍擊細胞培養皿側壁,使貼壁細胞震落。在超淨臺內加入腸上皮細胞培養液,並輕柔吹散細胞團塊。收集消化細胞,等分加入到新的細胞培養瓶中,並加入5ml細胞培養液,左右搖勻,做好標記,稍作靜置後放入到co2細胞培養箱中。上述腸上皮細胞培養液配方為加入了10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1×insulin-transferrin-selenium(胰島素-轉鐵蛋白-硒添加劑)的dmem培養基。若培養5代以後的細胞,上述培養基中的fbs含量可以調整為7.5%。
4、質粒轉染
在細胞6孔板中,接種106個細胞,待細胞長至80%的密度後,開始轉染實驗。分別250μl的opti-mem稀釋脂質體(lipofectamineltx)和總量5μg的sv40大t抗原質粒,均勻混合,室溫靜置10min。將兩種溶液均勻混合,室溫靜置20min,以便形成dna-脂質體複合物。將混合液體均勻加入培養皿中,輕輕搖晃培養皿混勻。37℃,5%co2培養6h,使用預熱好的pbs清洗,換新鮮培養液後繼續培養。待鋪滿細胞培養板後,正常傳代。
5、細胞的連續傳代培養
在細胞培養瓶中繼續培養已經轉染過sv40大t抗原質粒的細胞。在處於90%鋪滿狀態時進行傳代。在第5代後,將培養液中的血清含量調整為7.5%。細胞傳代過程中每隔20代對傳代細胞進行一次凍存備份,細胞繼續傳代至50代。
實施例2
1、細胞系的鑑定(轉錄組測序分析)
待培養至50代的兔小腸上皮細胞系生長至100%鋪滿時,加入含edta的0.25%胰酶消化液0.5ml,於含5%co2的37℃細胞培養箱中孵育5min,然後用吸管吹打使細胞脫落,加入培養基終止消化後,800g離心5min收集細胞,加入1mltrizol試劑(ambion,美國,貨號139605)。用試劑廠商提供的標準方法提取樣品總rna。
用分光光度計測定所提取的rna的濃度及純度。然後根據1%mops凝膠電泳(總rna上樣量約為1μg)結果,對提取的總rna進行rna完整性以及是否有基因組dna汙染的評價。
使用試劑盒ribo-zero消化核糖體rna後;加入打斷試劑將rna打斷成短片段,以打斷後的rna為模板,用六鹼基隨機引物合成一鏈cdna,然後配製二鏈合成反應體系合成二鏈cdna,在cdna二鏈合成時以dutp代替dttp,然後連接不同接頭,再利用ung酶法將含有dutp的一條鏈進行消化,只保留連接鏈不同接頭的cdna一鏈;使用試劑盒純化cdna一鏈;純化的cdna一鏈再進行末端修復、加a尾並連接測序接頭,然後進行片段大小選擇,最後進行pcr擴增;構建好的文庫用agilent2100bioanalyzer質檢合格後,使用illuminahiseqtm2500或其他測序儀進行測序。
經測序平臺測序所得的數據稱經過濾得到cleanreads,用tophat/bowtie2將cleanreads比對到兔基因組的參考序列。然後比對基因的轉錄情況。
發現該細胞系表達角蛋白8,e-降鈣素,ki67等腸上皮表面標誌分子,並且同時表達如lgr5,sox9等隱窩基底層幹細胞的表面分子。說明本細胞系保留了分化特徵。
2、永生化細胞生長速度測定
取第3代原代細胞與第50代傳代細胞1200個接種於24孔板內,使用5%的dmem培養液培養。每日對3孔細胞計數,求平均數後繪製生長曲線。生長曲線見圖3.
3、rt-pcr鑑定sv40大t抗原在兔小腸上皮細胞內的表達
提取轉染了永生化質粒後24h,48h後的第3代原代與第65代永生化兔小腸上皮細胞總rna並反轉錄為rna。具體方法參照下文。使用sv40大t抗原鑑定引物:上遊引物:actgcaaacaatggcctgag。下遊引物:taaaggcattccaccactgct。pcr反應條件為:預熱94℃,5min;變性94℃,30s;退火56℃,30s;延展72℃,30s;30個循環;最後延伸5min。使用百分之一瓊脂糖凝膠電泳,凝膠攝像系統拍照。由照片可得成功擴增出sv40大t抗原目的條帶,說明質粒可以在細胞內表達。電泳照片見圖4。
實施例3大型艾美耳球蟲體外培養
消化長滿一個25cm2細胞培養瓶的兔小腸上皮細胞,計數,並稀釋細胞。向細胞培養用6孔板中每孔加入106個細胞,並培養12h。待細胞貼壁之後,每孔均勻加入106個子孢子,繼續培養6h後。待細胞鋪滿後,使用加有5倍雙抗輕輕衝洗細胞培養板,衝洗掉沒入侵的子孢子,然後加入5%fbs,mem199培養基。培養換液時間設定為48h,94h。抽取出的舊培養液顯微鏡下鏡檢後,確保不存在逸出的裂殖子再棄去,如果存在裂殖子,離心後重新加入到新的細胞中繼續培養。在96h後,使用含有2.5%fbs的低血清mem199培養基繼續培養裂殖子。若細胞培養板細胞生長過滿,使用帶有edta的胰酶消化正常對細胞進行一傳二傳代。每天使用倒置螢光顯微鏡進行觀察。發現培養24h球蟲可發育到第一代裂殖體,第72h發育到第二代裂殖體,120h可以形成第三代滋養體。培養照片見圖5。
對比例1
將分離的隱窩細胞分別用培養基1(dmemf12,10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,50μg/ml的慶大黴素,1xinsulin-transferrin-selenium)。培養基2(dmemf12,10%fbs,表皮生長因子20μg/l,200iu/ml鏈黴素和青黴素,1xinsulin-transferrin-selenium)進行培養。發現使用培養基2進行培養,細胞在傳代2~3代後有明顯的細菌汙染。而使用培養基1細胞生長正常。
對比例2
分別連續傳代原代上皮細胞以及轉染的sv40大t基因的上皮細胞。可發現原代上皮細胞在第10代出現生長速度減緩,傳至15代後可見大部分細胞凋亡。而轉染sv40大t基因的上皮細胞可以不斷的在體外連續傳代,不存在生長停滯的現象。說明sv40大t基因可以永生化兔來源的小腸上皮細胞。
對比例3
分別將分離出來的隱窩細胞接種於質量比0.5%明膠包被和沒有包被的細胞培養板中。可發現使用明膠包被後的培養板中細胞團塊迅速貼壁並平鋪生長;而不使用明膠包被的培養板中細胞不能貼壁。
對比例4
在分離細胞的過程中,同時使用上述含有0.1~0.2mg/ml膠原酶和0.4~0.6mg/ml中性蛋白酶的消化液和質量比(m/m)2.5%的胰酶對兔小腸組織塊在37℃下進行消化。消化過程中不斷震蕩並使用顯微鏡隨時觀察,至隱窩細胞團大量釋放後終止消化。通過上述相差貼壁與重懸離心法提純的隱窩細胞團,並分別接種這兩種處理得到的隱窩細胞團於包被好明膠的細胞培養板內。可發現使用胰酶消化的隱窩細胞團塊不能良好的貼壁生長,而使用含有0.1~0.2mg/ml膠原酶和0.4~0.6mg/ml中性蛋白酶的消化液消化的隱窩細胞團塊是可以正常貼壁生長。
雖然,上文中已經用一般性說明及具體實施方案對本發明作了詳盡的描述,但在本發明基礎上,可以對之作一些修改或改進,這對本領域技術人員而言是顯而易見的。因此,在不偏離本發明精神的基礎上所做的這些修改或改進,均屬於本發明要求保護的範圍。