神經保護肽的製作方法
2023-05-16 13:22:56 3
本發明涉及具有神經保護活性的肽,所述肽用於治療中風和其他神經損傷或紊亂。本發明進一步涉及使用本發明的肽治療神經損傷或紊亂的方法。
背景技術:
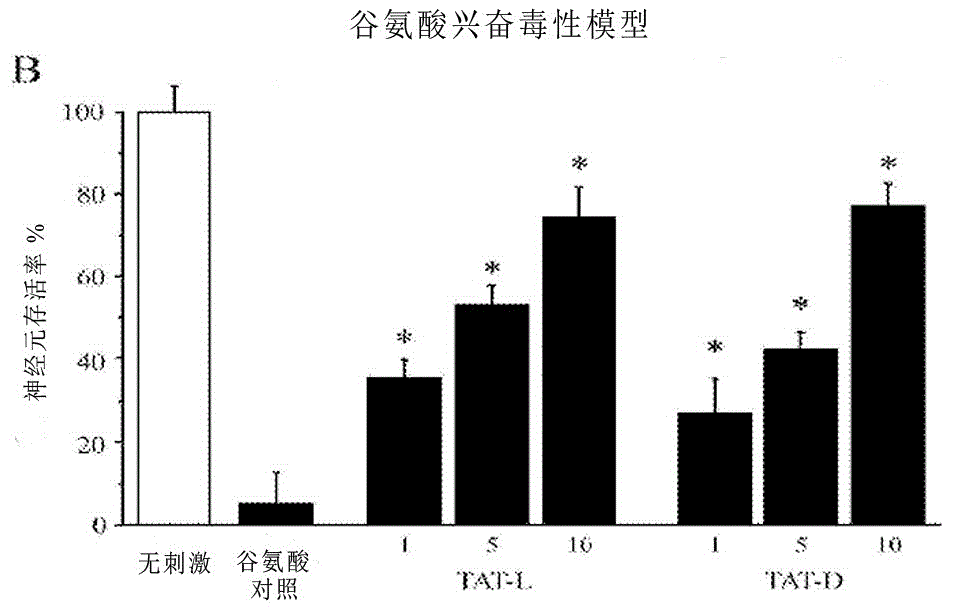
:細胞穿膜肽(CPPs)是小分子肽,用於促進正常不可滲透物質(比如其它肽、蛋白質、核酸或進入細胞的藥物)的傳遞。細胞穿膜肽(CPPs)的發展,也稱為肽轉導域蛋白(PTDs),自從在人類免疫缺陷病毒1型反式激活轉錄激活劑((Frankel和Pabo,1988;Green和Loewenstein,1988),通常被稱為TAT))中首次發現PTD,其作為治療藥物輸送的促進劑已經具有顯著進展。此後,這種序列的活性輸送部被分離出來(TAT48-58:稱為TAT肽),並且已經發現和合成超過100個新的CPPs(Milletti,2012)。融合CPPs的潛在療法已經在神經培養物系統和動物模型中進行了評估,模擬在各種紊亂中的神經損傷機制,包括腦缺血、癲癇、帕金森病和阿爾茨海默病(Lai等,2005;Liu等,2006;Arthur等,2007;Colombo等,2007;Nagel等,2008;Meade等,2009)。CPPs用於神經系統疾病是特別吸引人的,因為其具有穿過血腦屏障運輸物質,然後進入腦實質內的神經細胞的能力(Aarts等,2002;Zhang等,2013)。已經進入臨床試驗的CPP融合神經保護肽的兩個例子是JNK抑制劑肽(JNKI-1D-TAT或XG-102;ARAMIS,2012)和NMDA受體/突觸後密集區-95抑制肽(TAT-NR2B9c或NA-1;Dolgin,2012)。這兩種肽融合到TAT。任何CPP的一個重要特徵是臨床相關劑量的限制性毒性,並且非常需要限制性毒性的CPPs。同樣地,當治療神經損傷時非常需要神經保護性的肽。CPPs的結構和胺基酸含量差異很大,最近研究發現,在神經保護試驗中使用最廣泛的CPP—TAT肽也表現出具有本徵的神經保護性能。最近的研究(Xu等,2008;Vaslin等,2009;Meade等,2010a,b;Craig等,2011)已經報導在體外的興奮性毒性和糖氧剝奪,和在體內的腦室內注射後P12大鼠的腦局部缺血,TAT肽顯示了神經保護功效。雖然TAT的神經保護功效的確切機制尚不完全清楚,但有人猜測它幹擾NMDA受體激活(Xu等,2008;Vaslin等,2009),儘管一項研究未發現相互結合作用((Li等,2008)。此外,在使用CPP遞送構建體的RNAi研究中,在氣管內給藥後TAT和滲透肽兩者都單獨地顯示出下調肺內的MAP激酶mRNA(Moschos等,2007)。神經元或神經損傷或紊亂(比如偏頭痛、中風、創傷性腦損傷、脊髓損傷、癲癇)和神經變性疾病(包括亨廷頓氏病(HD)、帕金森氏病(PD)、阿爾茨海默氏病(AD)和肌萎縮性側索硬化症(ALS))是長期腦或脊髓損傷引起的發病和傷殘的主要原因。腦損傷通常涉及一系列細胞死亡過程,包括細胞凋亡、細胞自噬、壞死性凋亡和壞死,並且影響神經元星形膠質細胞、少突膠質細胞、小神經膠質細胞和血管內皮細胞(統稱為神經血管單元;NVU)。與神經損傷有關的破壞性觸發涉及包括穀氨酸興奮毒性鈣超載、氧化應激、蛋白水解酶和線粒體紊亂的多樣化途徑。如本文使用的術語「中風」包括任何影響大腦或脊髓的缺血性疾病,例如,腦或脊髓動脈的血栓閉塞、嚴重低血壓、圍產期缺氧缺血、心肌梗死、缺氧、腦出血、血管痙攣、外周血管病症、靜脈血栓形成、肺栓塞、短暫性腦缺血發作、肺缺血、不穩定型心絞痛、可逆性缺血性神經功能缺損、輔助溶栓活性、過度凝血條件、腦再灌注損傷、鐮狀細胞性貧血、中風紊亂或醫源性引起的缺血期,如血管成形術或腦缺血。通過由興奮性毒性引起的急性和延遲破壞處理,神經遞質穀氨酸的細胞外水平的增加可導致神經細胞死亡。細胞外穀氨酸的積累過度刺激NMDA和AMPA受體,導致細胞外鈣離子和鈉離子內流,以及蛋白結合鈣從細胞內儲存庫釋放。NMDA受體的過度激活還會觸發破壞分子(例如,一氧化氮、CLCA1;鈣激活的氯離子通道調節劑1、鈣蛋白酶、SREBP1︰固醇調節元件結合蛋白-1)和信號通路(例如DAPK;死亡相關的蛋白激酶,CamKII:鈣-鈣調蛋白依賴性蛋白激酶II)的產生。此外,穀氨酸誘導的神經元去極化和興奮性毒性通過電壓依賴性鈣通道(VGCC,例如CaV2.2,CaV3.3)、鈉鈣交換器(NCX)、酸敏感離子通道(ASIC)、瞬時受體電位陽離子通道2和7(TRPM2/7)和代謝型穀氨酸受體(mGluR),可以進一步觸發細胞內鈣離子流入。細胞內鈣離子的增加啟動一系列細胞損害事件,包括磷脂酶、蛋白酶、磷酸酶、激酶、NADPH氧化酶和一氧化氮合酶,並且啟動觸發細胞死亡的通路(即細胞凋亡、細胞自噬、程序性壞死(necroptosis)和壞死(necrosis))的激活。用於治療大腦局部缺血的神經退行性影響的化合物的例子,包括美國專利5,559,095,其描述了使用ω-芋螺毒素肽和結合併阻止電壓門控性鈣通道的相關的肽治療與神經元損傷有關的局部缺血的方法,和美國專利4,684,624,其描述了使用某些阿片肽的治療。進一步的例子包括美國專利申請公開2009/0281036,公開了連接其他肽的融合肽的使用,通過抑制NMDA受體與NMDAR相互作用蛋白的相互作用,以減少對哺乳動物細胞損傷的破壞性影響。同樣,US2012/0027781公開了連接靶向肽和其他肽的使用,以提供神經保護功能。US6,251,854公開了一種提供對抗興奮毒性的神經元損傷的保護的化合物,其選自結合結構式1化合物的短富含精氨酸的寡肽︰許多潛在的神經保護劑也在低至中劑量時表現出毒性。許多CPPs也在低至中劑量表現出毒性。因此,需要神經保護肽,其在低劑量有效,具有低細胞毒性並且其能提供對抗超過一種類型的神經損傷的保護。這裡討論的
背景技術:
旨在便於對本發明的理解。並非意圖承認或認可提及的任何材料是或曾是本申請優先權日的公知常識的一部分。技術實現要素:根據本發明的一個方面,提供一種分離的、鹼性(即陽離子)富含胺基酸的肽作為神經保護劑的應用。這種分離的肽可以是CPP。因此,本發明延伸至具有神經保護活性的分離的、陽離子富含胺基酸的肽。本發明延伸至具有神經保護活性的肽的功能性片段。在pH值為7時,所述肽的淨電荷可以是8或更高,優選地,在pH值為7時,淨電荷是10或更高的,更優選地在pH值為7時,淨電荷是11或更高。所述肽可以是非天然存在的肽。因此,本發明的肽可以是人造肽。形成的肽的一部分的陽離子(即鹼性)胺基酸殘基可以是精氨酸或賴氨酸。因此,所述肽可以富含精氨酸。作為替換,或另外的,所述肽可以富含賴氨酸,和/或富含色氨酸。所述肽的長度可以在10個胺基酸和100個胺基酸之間,優選地在10個胺基酸和32個胺基酸之間。所述肽可以具有序列:X(K)-Z(N)-X(K)-Z(N)-X(K)其中X可以是任何天然存在或合成的胺基酸,包括陽離子(即鹼性)胺基酸殘基;K是1和5之間的整數;Z是鹼性(即陽離子)胺基酸殘基;和N是1和30之間的整數。胺基酸取代「X」位置,優選不會降低神經保護肽的神經保護功效的胺基酸。在一個實施例中,Z可以是精氨酸。在另一個實施例中,Z可以是賴氨酸。這種肽可以包含至少一個連續的富含精氨酸片段。在其他實施例中,該肽可以包含非連續的富含精氨酸片段。所包含的鹼性(即陽離子)胺基酸殘基的比例為佔肽或片段的至少30%,優選地,至少40%,優選地,至少50%,更優選地,至少60%,在某些情況下高達肽或片段的100%。所述肽含有精氨酸的含量可以超過肽中胺基酸含量的20%,優選超過30%、超過40%、超過50%、超過60%、超過70%、超過80%、超過90%、超過95%、超過99%,或更優選100%。在本發明的某些實施例中,肽可以含有聯合的精氨酸和賴氨酸的含量超過40%、超過50%、超過60%、超過70%、超過80%、超過90%、超過95%、超過99%,或更優選100%。所述肽可以包括多個單陽離子胺基酸殘基,比如精氨酸殘基,被其他胺基酸殘基穿插,尤其是,可以被鹼性(即陽離子)胺基酸殘基穿插,如賴氨酸,和/或色氨酸。所述肽可以包括在相鄰位置上精氨酸殘基的重複序列,如RR,或RRR,或RRRRR,或更高階的重複,和可以穿插在其他胺基酸之間或胺基酸的區段之間。因此,所述肽可以完全由陽離子胺基酸組成。在一個實施例中,該肽完全由精氨酸殘基組成。根據本發明的一個方面,提供了一種長度為10至32個胺基酸殘基的分離的肽用於治療神經損傷的應用,其中該分離的肽包括至少10至22個殘基。在本發明的優選實施例中,該肽可以是用於治療神經損傷的長度為10至32個胺基酸殘基的分離的肽,其中該分離的肽包含至少10至22個的精氨酸殘基。所述分離的肽具有的精氨酸殘基含量為至少30%、至少40%、至少50%、至少60%、至少70%、至少80%、至少90%,或至少99%。所述分離的肽可以是包含10至18個精氨酸殘基的聚精氨酸肽。在優選的實施例中,該分離的肽是R12、R15或R18,更優選R15。具體來說,所述分離的肽可包含任何一種或多種選自由序列號(SEQ.ID.NOS.)為SEQ.ID.NOS.7、8、9、10、11、12、13、16、30、31、32、33、34、35、36、37以及它們的具有神經保護活性的功能性片段和其類似物組成的組中的肽。所述分離的肽可包括至少一個包括至少4個連續的精氨酸殘基的聚精氨酸片段,優選至少兩個聚精氨酸段,更優選地三個聚精氨酸段。所述分離的多肽可以是穿膜肽((SEQ.ID.NO.22)。所述分離的肽可以包含魚精蛋白衍生物的混合物。魚精蛋白衍生物的混合物可包含硫酸魚精蛋白。所述分離的肽可以影響細胞的內吞過程;影響細胞表面受體的功能,實現減少細胞鈣內流;與線粒體外膜相互作用和/或穩定線粒體外膜以維持線粒體的功能;或抑制、下調或影響鈣依賴性前蛋白轉化酶弗林蛋白酶。所述分離的肽可以是合成肽或人造肽。該分離的肽可以被包括在或融合到其他多肽中。該分離的多肽可以與至少一個接頭序列彼此融合。所述分離的肽可以在小於50μM的IC50水平上表現出神經保護活性,優選小於20μM,更優選小於10μM。本發明延伸到分離的肽製備用於治療或預防神經損傷的藥物組合物或藥物中的應用,所述分離的肽包含任何一種或多種序列號(SEQ.ID.NOS.)為SEQ.ID.NOS.7、8、9、10、11、12、13、16、30、31、32、33、34、35、36、37,以及它們的具有神經保護活性的功能性片段及其類似物。所述藥物組合物或藥物可以用於治療或預防神經損傷,該藥物組合物或藥物包括本發明的分離的肽或本發明的多核苷酸序列。所述藥物組合物或藥物可以用於治療或者預防局部缺血、圍產期缺氧缺血、阿爾茨海默氏病、亨廷頓氏病、多發性硬化症、帕金森氏症、肌萎縮性側索硬化、中風、外周神經病、脊髓損傷或癲癇。所述藥物組合物或藥物可以包含藥學上可接受的載體、輔助劑或媒介物。根據本發明的另一個方面,提供了一種治療神經損傷或促進神經元存活的方法,該方法包括給有需要的患者施用藥學上可接受的和/或藥學有效量的本發明的肽或本發明的藥物組合物或藥物的步驟。根據本發明的又一個方面,提供了一種抑制受試者的神經元細胞死亡的方法,包括給需要這種治療的受試者施用藥學上可接受的和/或藥學有效量的本發明的肽或者本發明的藥物組合物或藥物。給藥劑量可以從0.001mg/kg至50mg/kg。本發明延伸到試劑盒,其包括在一個或多個容器中的本發明的藥物組合物或藥物以及關於用法說明的使用說明書或信息手冊和/或關於藥物組合物的使用的信息。更具體來說,所述肽可以是任何一種或多種具有序列號(SEQ.ID.NOS.)為SEQ.ID.NOS.7、8、9、10、11、12、13、16、30、31、32、33、34、35、36、37的序列的肽,或者可以包括具有與所述肽至少60%、優選70%、更優選80%、較優選90%、更優選99%或更高的序列同一性的序列,這些肽具有神經保護活性。所述肽可以包含在R8和R22之間任何長度的聚精氨酸肽,或其重複序列。在一個實施例中,所述肽選自包含R8、R9、R10、R11、R12、R13、R14、R15、R16、R17、和R18的組。在一個優選的實施例中,肽是R10。在另一個優選的實施例中,肽是R12。在又一個優選的實施例中,所述肽為R15。在另一個優選的實施方案中,肽是R18。在神經損傷的治療或預防中,肽的應用可以包括任何兩種或多種本發明的肽的混合物的應用,特別是SEQ.ID.NOs32至36。在另一個實施例中,所述應用可以包括在神經損傷的治療或預防中,使用SEQ.ID.NO.37的肽,或者SEQ.ID.NO.37與任意SEQ.ID.NOS32至36的肽一起使用。在本發明的另一個方面,提供SEQ.ID.NOs32至37的分離肽,包括與所述肽具有至少60%、70%、80%、90%、甚至99%的序列或更高的序列同一性的序列,這些肽具有神經保護活性。所述肽可以是市售的從鮭魚精分離的精蛋白肽混合物。因此,所述肽可以是以包括硫酸魚精蛋白的混合物的形式,如歐洲藥品管理局(EuropeanMedicinesAgency)所描述的「AssessmentReportforProtaminecontainingmedicinalproducts」(含有藥品的魚精蛋白評估報告),在2012年11月15日公布的編號為EMEA/H/A-5(3)/1341,其內容僅以參考方式併入本文。表1顯示了不同形式的鮭魚精子魚精蛋白的概述。魚精蛋白肽序列(鮭魚)存在於用於臨床使用的硫酸魚精蛋白中或來自SwissProt資料庫。如本說明書中所使用的,術語「魚精蛋白」指的是市售的可注射形式或硫酸魚精蛋白,其包括SEQ.ID.NOs32至35的肽的混合物。所述肽可以以如下百分比例存在於肽的混合物中:在優選的實施例中,肽是以如下百分比例存在於肽的混合物中:SEQ.ID.NO標識符百分比32Ptm118–2233Ptm230–3534Ptm321.5–2835Ptm419–23在更優選的實施例中,肽是以如下百分比例存在於肽的混合物中:SEQ.ID.NO標識符百分比32Ptm120.133Ptm233.534Ptm325.135Ptm421.3更具體地,所述肽的混合物可以是SEQ.ID.NOs32、33、34和35的肽的混合物,市售可得的是硫酸魚精蛋白(鮭魚),如SanofiAventis生產的。因此,肽的混合物可以與氯化鈉、鹽酸、氫氧化鈉和水混合。肽的混合物可以在可注射或靜脈給藥的輸送製劑中。肽存在於輸送製劑中的濃度範圍可以為0.1mg/mL至100mg/mL,優選1mg/mL至20mg/mL,更優選10mg/mL。在使用中,當為可注射或靜脈注射的製劑的形式時,輸送製劑可以通過在1至30分鐘的時間內緩慢靜脈注射給藥,優選5至20分鐘,更優選超過10分鐘的時間。所述肽可以具有細胞穿透活性。因此,所述肽可以是CPP。所述肽可以包括在相鄰位置上重複的精氨酸殘基,例如RR,或RRR,或RRRR,或更高階的重複,並且可能穿插在其他胺基酸之間,或胺基酸的區段之間。根據本發明的另一方面,提供本發明的肽在製備用於治療神經損傷的藥物組合物或藥物中應用。本發明延伸到本發明的肽混合物在製備用於治療神經損傷的藥物中應用。根據本發明的再一個方面,提供了使用本發明的肽或藥物組合物影響與鈣離子內流相關聯,更具體地說與NMDA、AMPA、VGCCs、NCX、TRMP2/7、ASIC和mGlu受體相互作用,更具體地說還使得能夠減少細胞外鈣離子內流相關聯,的細胞表面受體的功能。在本發明的另一實施例中,提供了使用本發明的肽或藥物組合物與線粒體外膜相互作用和/或穩定線粒體外膜,從而有助於保持線粒體功能。根據本發明的又一個方面,提供了使用本發明的肽或藥物組合物,用於抑制、下調或影響鈣依賴性前蛋白轉化酶弗林蛋白酶。根據本發明的另一個方面,提供了用於治療神經損傷的藥物組合物或藥物,所述藥物組合物或藥物包括本發明的分離的肽,或任意一種或多種本發明的分離的肽。藥物組合物或藥物可以包括藥學上可接受的載體、輔助劑或媒介物。根據本發明的另一個方面,提供了一種治療神經損傷的方法,該方法包括給有需要的患者施用藥學上可接受的和/或藥物有效量的本發明的肽的步驟。患者可以施用生理學上可接受量的本發明的肽。根據本發明的再一個方面,提供了一種抑制受試者神經元細胞死亡的方法,該方法包括給需要這種治療的受試者施用有效量的神經保護性肽,以抑制受試者的神經元細胞死亡,所述神經保護性肽是任何一種或多種本發明的肽。患者可以施用生理學上可接受量的任何一種或多種本發明的肽的混合物。更具體地,患者可以施用一種或多種選自由SEQ.ID.NOs32、33、34、35、36或37組成的組中的肽的混合物。在本發明的一個特定方面,患者可以施用包含SEQ.ID.NOs32、33、34和35的混合物。在其它組合中,患者可以施用選自由SEQ.ID.NOs32、33、34和35組成組中的肽與SEQ.ID.NO.36的肽的任意組合。在一個具體的實施例中,患者可以施用SEQ.ID.NO.35。本發明的的肽可以提供抗神經遞質受體拮抗劑的神經保護活性。神經遞質受體可以是受NMDA、穀氨酸鹽、紅藻氨酸或缺血性過程約束、相互作用或影響的受體。本發明的肽可以包括在其他多肽中或可以融合到其他多肽中。本發明的肽可連接或融合在這些其他多肽的N-或C-末端。本發明的肽可連接或融合到這些其他多肽,以使本發明的肽存在於適於治療神經損傷的構型中。作為替換,或者另外的,一種或多種本發明的肽可以與接頭序列相互融合。所述接頭序列可以包括任何胺基酸序列,包括但不是限於鹼性/陽離子富含胺基酸的接頭序列。作為替換,或者另外的,所述接頭序列可以是可裂解的接頭序列。在一個實施例中,所述接頭可以是一個或多個MMP型接頭、鈣蛋白酶、半胱天冬蛋白酶或tPA接頭。MMP型接頭被定義為通過基質金屬蛋白酶(MMPs)識別和裂解的肽序列。同樣地,鈣蛋白酶、半胱天冬蛋白酶或tPA接頭是被這些蛋白酶(即分別是鈣蛋白酶、半胱天冬蛋白酶或tPA)的識別和裂解的肽序列。本發明的肽也可以融合到其他肽,被運送到神經損傷的位點或經細胞內運輸進入神經細胞或在神經細胞內。本發明的肽也可以連接到輔助肽,所述輔助肽可以與對神經功能有害的酶結合或者相互作用,由此輔助肽可以作為跨過細胞膜運輸後酶的競爭性抑制劑。輔助肽可以選自tPA、鈣蛋白酶和基質金屬蛋白酶,並且可以是使用可切割接頭(比如半胱天冬蛋白酶序列)連接到本發明的肽,使得tPA、鈣蛋白酶或基質金屬蛋白酶可以從本發明的肽中釋放出來,然後可以作為對神經功能有害的酶的細胞內競爭性抑制劑。因此,本發明延伸至連接到半胱天冬蛋白酶裂解位點的本發明的肽,它本身又與鈣蛋白酶、tPA或基質金屬蛋白酶連接。本發明的肽可以在小於10μM的IC50水平表現出神經保護活性,優選小於5μM,優選小於1μM,在某些情況下低至或低於0.2μM,甚至低至0.1μM。如上文提到的,所述肽可以包括本發明肽序列的重複序列,或其功能性片段,即表現出神經保護活性的片段。因此,本發明的一個方面提供了一種分離的多肽,具有包含在其中的表現出神經保護功效的肽段,所述肽段的長度為8至100個之間胺基酸殘基,其中該神經保護肽選自具有鹼性/陽離子胺基酸含量超過所述肽段長度20%的肽段,優選超過30%、超過40%、超過50%、超過60%、超過70%、超過80%以上、超過90%、超過95%、超過99%或更優選地100%。根據本發明的另一個方面,提供了編碼本發明的肽的分離的多核苷酸序列,與分離的多核苷酸序列互補的序列,以及與所述分離的多核苷酸序列具有至少60%、70%、80%、90%、95%、99%或者100%的同源性的序列。多核苷酸序列可以是一種或多種的分離序列並且可以是在嚴格條件下與本發明的多核苷酸序列雜交的序列。所述多核苷酸序列可以是非天然存在的多核苷酸序列或DNA。因此,它們可以包括人造的、人工構建體,比如cDNA。本發明中包括載體,比如表達載體,其包括分離的上述分離核酸以及用這些載體或DNA序列轉化的細胞。多核苷酸序列可以編碼魚精蛋白。多核苷酸序列可以是SEQ.ID.NO.38的序列。在本發明的另一個方面,提供了選自由SEQ.ID.NOS.7、8、9、10、11、12、13、16、30、31、32、33、34、35、36和37組成的組中的分離的肽,其包括與所述肽具有至少60%、優選地70%、更優選地80%、甚至更優選地90%、更更優選地99%或更高的序列同一性的序列,所述的肽具有神經保護活性。也提供了在藥物製劑或藥物的製備中所述分離的核酸、載體或細胞的應用。此外,本發明提供了試劑盒,其包括在上述製劑的至少一種中(在一個或多個容器中)的上述藥物組合物和關於用法說明的使用說明書或信息手冊和/或關於所述藥物組合物的使用的信息。在一個實施例中,藥物組合物包括如上所限定的本發明的肽,其用於治療局部缺血、阿爾茨海默氏病、帕金森氏症、肌萎縮性側索硬化、中風、外周神經病、脊髓損傷或癲癇。在另一個實施例中,本發明提供了一種促進神經元的存活的方法,該方法包括將本發明的肽或其組合物與神經元接觸的步驟。優選地,該方法在體外進行。在其另一個方面,本發明提供了一種保護血腦屏障的內皮細胞免受OGD或局部缺血的方法和組合物。這些發現證明本發明的肽具有用於抑制或改善與興奮性中毒和缺血性損傷相關的神經損傷事件/途徑的能力,或者可以在用於抑制或改善與興奮性中毒和缺血性損傷相關的神經損傷事件/途徑的方法中使用。另外,如本文所包含的在預刺激暴露試驗(pre-insultexposuretrial)中魚精蛋白和魚精蛋白衍生物的作用所示,一個新的重要發現是,神經元在暴露於穀氨酸或OGD前的1至4小時採用魚精蛋白治療,可以通過減少細胞死亡引發神經保護反應。這點很重要,因為在許多腦血管(例如頸動脈內膜切除術)和心血管(如冠狀動脈旁路移植)的外科手術中,患者有遭受腦缺血或中風導致腦損傷的風險。因此,本發明的方法延伸到在這些手術前的0.25小時至4小時,優選0.5至3小時,更優選1至2小時的時間窗口施用至少一種本發明的肽、藥物或藥物組合物,以保護大腦免受任何此類腦缺血事件。因此,本發明延伸到使用至少一種本發明的肽用於治療或預防神經損傷、腦血管損害或損傷、心血管損害或損傷,或用於病人可能有患有腦缺血或中風的風險的外科手術中。本文所公開的本發明的序列的初級胺基酸序列的微小修飾都可能導致蛋白質具有實質上相同或增強的活性。這些修飾可以是特意的,如通過定向誘變,或者可以是偶然的,例如通過生產魚精蛋白或LMWP的生物體的宿主突變。所有這些修飾都包括在本發明的範圍內,只要保有神經保護活性。附圖說明本發明的進一步特徵通過以下幾個僅用於舉例說明本發明的目的的非限制性實施例進行全面的描述。以下的描述不是對如上所述的本發明的概括性總結、公開內容或描述的限制,並且參照附圖,以下的描述中的術語「魚精蛋白」或「Ptm」指的是市售的硫酸魚精蛋白(由幾個製造商製造,包括SanofiAventis,尤其指定Hoffman,1990年),而「Ptm1」到「Ptm」5分別指SEQ.ID.NOs32、33、34、35和36,並且其中:圖1a顯示了穀氨酸興奮毒性模型的結果;肽的濃度以μM表示。A:神經元暴露於穀氨酸並採用CPPs、陽性對照肽(JNKI-1D-TAT/PYC36L-TAT)和穀氨酸受體阻滯劑(Blkrs;5μM:MK801/5μM:CNQX)處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4-6;*P<0.05)。圖1b顯示了穀氨酸興奮毒性模型的進一步結果;肽的濃度以μM表示。神經元暴露於穀氨酸並採用TAT-L和TAT-D肽處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4-6;*p<0.05)。圖1c顯示了穀氨酸興奮毒性模型的進一步結果。具體地說,當在穀氨酸刺激之前衝淨肽時肽的功效;肽的濃度以μM表示。當在刺激之前衝淨CPPs時,神經元暴露於穀氨酸後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4-6;*P<0.05)。圖1d顯示了穀氨酸興奮毒性模型的進一步結果。當在穀氨酸刺激後添加肽時肽的功效。當在刺激後第0或15分鐘添加Arg-9和Arg-12肽以及對照肽JNKI-1D-TAT和NR29c時,神經元暴露於穀氨酸後24小時的神經元存活率;在這個實驗中,暴露於穀氨酸的對照組中導致的細胞死亡少於其它實驗組(60%對95%)。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4-6;*P<0.05)。圖1e顯示了穀氨酸興奮毒性模型的進一步結果;肽的濃度以μM表示。A:神經元暴露於穀氨酸和用R9、R12、R15、R18和R9/tPA/R9處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖1f顯示了穀氨酸興奮毒性模型的進一步結果;肽的濃度以μM表示。神經元暴露於穀氨酸和用R9、E9/R9、DAHK和PTD4處理後24小時的神經元存活率。MTS數據表示為在沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖1g顯示了穀氨酸興奮毒性模型的進一步結果;肽的濃度以μM表示。神經元暴露於穀氨酸和用R9和NR29c和對照肽PCY36(PYC36L-TAT)處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖1h顯示了穀氨酸興奮毒性模型的進一步結果:溫和刺激;肽的濃度以μM表示。神經元暴露於穀氨酸和用R9、R12和NR29c和對照肽JNK(JNKI-1-TAT)處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖1i顯示了穀氨酸興奮毒性模型的進一步結果:溫和刺激;肽的濃度以μM表示。神經元暴露於穀氨酸和用R1、R3、R6、R9、R12和NR29c對照肽處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖1j顯示了穀氨酸興奮毒性模型的進一步結果;肽濃度=5μM。神經元暴露於穀氨酸和用R1、R3、舊R9(邁拓)、新R9(中肽生化有限公司)、R12處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖2顯示了紅藻氨酸興奮毒性模型的結果;肽的濃度以μM表示。神經元暴露於紅藻氨酸和用CPPs、陽性對照肽(JNKI-1D-TAT/PYC36L-TAT)和穀氨酸受體阻滯劑(Blkrs;5μM:MK801/5μM:CNQX)處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖3a顯示了體外局部缺血模型的結果。肽出現在體外局部缺血期間且在局部缺血後為50%劑量:肽的濃度以μM表示。在體外局部缺血和用CPPs、陽性對照肽(PYC36L-TAT)和穀氨酸受體阻滯劑(Blkrs;5μM:MK801/5μM:CNQX)處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖3b顯示了體外局部缺血模型的進一步結果。肽出現在體外局部缺血後:肽的濃度以μM表示。在體外局部缺血和用R9、R12、R15和R18處理後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SEM;n=4;*P<0.05)。圖3c顯示了體外局部缺血模型的進一步結果。R9肽出現在體外局部缺血期間且在局部缺血後為50%劑量:bEND3細胞的肽劑量在體外局部缺血期間為10μM/在體外局部缺血後為5μM。SH-5YSY細胞的肽劑量在體外局部缺血期間為5μM/在體外局部缺血後為2.5μM。體外局部缺血後24小時的細胞存活率(MTS數據均值±SEM;n=4;*P<0.05)。圖4顯示了培養物暴露於不同肽濃度後的神經元存活率。肽濃度以μM表示。暴露於R3、R6、R9、R12、R15、R18、JNK(JNKI-1D-TAT)和NR29c肽後24小時的神經元存活率。細胞存活率數據表示為490nm處的MTS吸光度(均值±SEM;n=4)。圖5顯示了初始動物模型預試驗的結果,顯示了R9D肽在大鼠永久中腦動脈阻塞(MCAO)中風模型中的功效。R9D肽在MCAO後30分鐘靜脈注射給藥。梗死體積(腦損傷)在MCAO後24小時測量(均值±SD)。圖6顯示了在穀氨酸鹽模型中使用R9、R10、R11、R12、R13和R14的劑量響應研究。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。圖7顯示了在穀氨酸鹽模型中使用R9D、R13、R14和R15的劑量響應研究。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。圖8顯示了在穀氨酸鹽模型中使用R6、R7、R8和R9的劑量響應研究。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。圖9顯示了穀氨酸興奮毒性模型的結果;魚精蛋白濃度以μM表示。在暴露於穀氨酸(100μM)前採用魚精蛋白處理神經元培養物15分鐘,然後在37℃下暴露於穀氨酸(100μM)5分鐘。暴露於穀氨酸和用不同魚精蛋白濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖10顯示了穀氨酸興奮毒性模型的結果;魚精蛋白濃度以μM表示。在暴露於穀氨酸(100μM)前和暴露於穀氨酸的過程中,採用魚精蛋白處理神經元培養物15分鐘,然後在37℃下暴露於穀氨酸5分鐘。暴露於穀氨酸和用不同魚精蛋白濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖11顯示了穀氨酸興奮毒性模型的結果;魚精蛋白濃度以μM表示。神經元培養物暴露於穀氨酸(100μM)(在37℃下進行5分鐘),僅在此之前採用魚精蛋白處理神經元培養物5或10分鐘。暴露於穀氨酸和用不同魚精蛋白濃度處理或不處理(穀氨酸)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖12顯示了穀氨酸興奮毒性模型的結果;魚精蛋白和低分子量魚精蛋白(LMWP)的濃度以μM表示。神經元培養物暴露於穀氨酸(100μM)在37℃下進行5分鐘,在此之前及其期間採用魚精蛋白或LMWP處理神經元培養物15分鐘。暴露於穀氨酸和用不同魚精蛋白或LMWP濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖13顯示了穀氨酸興奮毒性模型的結果;魚精蛋白和低分子量魚精蛋白(LMWP)的濃度以μM表示。神經元培養物暴露於穀氨酸(100μM)在37℃下進行5分鐘,在此之前及其期間採用魚精蛋白或LMWP處理神經元培養物15分鐘。暴露於穀氨酸和用不同魚精蛋白或LMWP濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖14顯示了穀氨酸興奮毒性模型的結果;魚精蛋白和LMWP濃度以μM表示。神經元培養物暴露於穀氨酸(100μM;在37℃下進行5分鐘),僅在此之前立即或在此之前1或2小時,用魚精蛋白處理神經元培養物10分鐘。在暴露於穀氨酸和用不同魚精蛋白或LMWP濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖15顯示了穀氨酸興奮毒性模型的結果;魚精蛋白和魚精蛋白肽1至5(「Ptm1至5」)的濃度以μM表示。神經元培養物暴露於穀氨酸(100μM)在37℃下進行5分鐘,在此之前用魚精蛋白和魚精蛋白肽處理神經元培養物15分鐘。暴露於穀氨酸和用不同魚精蛋白和魚精蛋白肽濃度處理或不處理(穀氨酸對照)後24小時的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖16顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白濃度以μM表示。魚精蛋白處理神經元培養物在氧糖剝奪(OGD)進行50分鐘後立即進行並直到試驗結束(24小時)。OGD和用不同魚精蛋白濃度處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖17顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白濃度以μM表示。採用魚精蛋白處理神經元培養物在OGD進行50分鐘後立即進行並直到試驗結束(24小時)。OGD和用不同魚精蛋白濃度處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖18顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白和LMWP濃度以μM表示。在OGD進行50分鐘前1小時,用魚精蛋白或LMWP處理神經元培養物10分鐘。OGD和用不同的魚精蛋白濃度預處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖19顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白濃度以μM表示。在OGD進行50分鐘後,採用魚精蛋白處理神經元培養物15或30分鐘。OGD和用不同魚精蛋白濃度後處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖20顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白和LMWP的濃度以μM表示。在OGD進行50分鐘後,採用魚精蛋白或LMWP處理神經元培養物15分鐘。OGD和用不同魚精蛋白或LMWP濃度後處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖21顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白和魚精蛋白肽的濃度以μM(5μM)表示。在OGD進行45分鐘後,採用魚精蛋白處理神經元培養物15分鐘。OGD和用不同魚精蛋白或魚精蛋白肽濃度後處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=5;*P<0.05)。圖22顯示了氧糖剝奪(OGD)模型的結果;魚精蛋白、魚精蛋白和低分子量魚精蛋白(LMWP)肽的濃度以μM(5μM)表示。在OGD進行45分鐘後,採用魚精蛋白處理神經元培養物15分鐘。OGD和用不同魚精蛋白或魚精蛋白肽濃度後處理或不處理(OGD)後24小時的神經元存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖23顯示了使用腦內皮細胞(bEND3細胞)的氧糖剝奪(OGD)模型的結果;魚精蛋白的濃度以μM表示。就在3個不同OGD期間(2h15min、2h30min和2h45min)之前且直到試驗結束(24h),採用魚精蛋白處理bEND3培養物15分鐘。測定OGD後24小時的細胞存活率。MTS數據表示為在以沒有OGD對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖24顯示了魚精蛋白肽暴露於腦內皮細胞(bEND3細胞)的結果;魚精蛋白濃度為1μM。就在OGD(2h30min的持續時間)前,將魚精蛋白肽1-5(Ptm1-Ptm5)、硫酸魚精蛋白(Ptm)和低分子量魚精蛋白(LMWP)添加到bEND3細胞培養物中15分鐘。通過MTS法評估OGO後24小時的細胞存活率。MTS數據表示為490mm處的吸光度值(均值±SD;n=4-6;*P<0.05)。圖25顯示了魚精蛋白暴露於腦內皮細胞(bEND3細胞)的結果;魚精蛋白的濃度以μM表示。採用魚精蛋白處理bEND3培養物0.5、1或2小時。細胞暴露於魚精蛋白或不處理(對照)後2小時的細胞存活率。MTS數據表示為在490mm處的吸光度值(均值±SD;n=4-6;*P<0.05)。圖26顯示了魚精蛋白暴露於腦內皮細胞(bEND3細胞)的結果;魚精蛋白的濃度以μM表示。採用魚精蛋白處理bEND3培養物2小時。細胞暴露於魚精蛋白或不處理(對照)後2小時的細胞存活率。MTS數據表示為在490mm處的吸光度值(均值±SD;n=4-6;*P<0.05)。圖27顯示了在永久中腦動脈阻塞(MCAO)中風模型中,閉塞後30分鐘靜脈注射R12、R15、R18和魚精蛋白(Ptm)肽的神經保護功效。肽劑量為1μmol/kg(600μl:IV)且MCAO後24小時進行梗死評估(均值±SE;n=8-10;*P<0.05)。動物處理是隨機的且所有的步驟都採用盲法進行。圖28顯示了當僅在暴露於穀氨酸的5分鐘期間肽存在於神經元培養物中時,R9、R12、R15和R18在穀氨酸興奮毒性模型中的神經保護功效。測定暴露於穀氨酸後20-24小時的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖29顯示了肽僅在暴露於穀氨酸前存在於神經元培養物中10分鐘時,R9、R12、R15和R18在穀氨酸模型中的神經保護功效。穀氨酸後20-24小時測定神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖30顯示了僅在暴露於穀氨酸前1-5h時肽存在於神經元培養物中10分鐘時,R12和R15在穀氨酸模型中的神經保護功效。穀氨酸後20-24小時測定神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的其情況下神經元存活的百分比(均值±SD;N=4;*P<0.05)。圖31顯示了在糖氧剝奪(OGD)模型中,當肽在OGD後立刻添加到神經元培養物中並且在15分鐘後移除時,R9、R12、R15和R18的神經保護功效。OGD後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖32中顯示了當在OGD前1-3h肽存在於神經元培養物中僅10分鐘時,R9、R12、R15和R18在糖氧剝奪中的神經保護功效。OGD後20-24小時測量的神經元存活率。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4-6;*P<0.05)。圖33顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物中15分鐘時,PTD4、E9/R9和R9在穀氨酸興奮毒性模型中的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖34顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物15中分鐘時,XIP和PYC36-TAT在穀氨酸興奮毒性模型中的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖35顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物中15分鐘,NCXBP3在穀氨酸興奮毒性模型中的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖36顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物中15分鐘,K10、R10和TAT-NR2B9c在穀氨酸興奮毒性模型中的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖37顯示了在暴露於NMDA前和暴露於NMDA經過5分鐘期間,肽存在於神經元培養物中15分鐘時,R15和TAT-NR2B9c在NMDA興奮毒性模型中的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖38顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物中15分鐘,R8和Cal/R9在穀氨酸興奮毒性模型中的神經保護功效。穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖39顯示了僅在暴露於穀氨酸前,R9D、R12、R15和PYC36-TAT肽添加到神經元培養物中10分鐘前,R9D、R12、R15和PYC36-TAT肽用±肝素(20IU/ml)在室溫下培養5分鐘後其神經保護功效。穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖40顯示了在暴露於穀氨酸前,神經元培養物以±肝素(40IU/ml)在37℃下培養5分鐘,然後添加肽僅在37℃下培養10分鐘然後移除時,R9D、R12、R15和穀氨酸受體阻滯劑(Blks︰5μMMK801/5μMCNQX)的神經保護功效。暴露於穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖41顯示了在暴露於穀氨酸前和暴露於穀氨酸5分鐘期間,肽存在於神經元培養物中15分鐘,kFGF、kFGF-JNKI-1、TAT-JNKI-1和JNKI--1TATD在穀氨酸興奮毒性模型中的神經保護功效。穀氨酸後20-24小時測定的神經元存活率。肽濃度以μM表示。MTS數據表示為在以沒有刺激對照作為100%存活率的情況下神經元存活的百分比(均值±SD;n=4;*P<0.05)。圖42顯示了在氧糖剝奪前,硫酸魚精蛋白(Ptm)和R18肽預暴露原代鼠星形膠質細胞15分鐘的結果。肽濃度為2μM。通過MTS法評價OGD後24小時的細胞存活率。MTS數據表示為在490mm處的吸光度值(均值±SD;n=4-6;*P<0.05)。圖43顯示了原代大鼠星形膠質細胞暴露於魚精蛋白肽1-5(Ptm1-Ptm5)、硫酸魚精蛋白(Ptm)和低分子量魚精蛋白(LMWP)的結果;魚精蛋白的濃度以μM表示。用魚精蛋白處理星形膠質細胞24小時。暴露於魚精蛋白或不處理(對照)後24小時的細胞存活率。MTS數據表示為在490mm處的吸光度值(n=4)。圖44顯示了神經元培養物暴露於穀氨酸後,穀氨酸受體阻滯劑(MK801/CNQX以5μM/5μM)、R9D、R15、PYC36-TAT、TAT、TAT-NR2B9cJNKI-1-TATD、TAT-JNKI-1、kFGF-JNKI-1和kFGF的細胞內鈣離子內流動力學。Fura-2AM用於細胞內鈣評估。代表螢光Fura-2AM示蹤劑;添加(箭頭)穀氨酸(100μM終濃度)前30秒和添加(箭頭)穀氨酸(100μM終濃度)後的神經元培養物的螢光強度(FI)。肽或穀氨酸受體阻滯劑添加到神經元培養物反應10分鐘並且在穀氨酸前移除。數值是均值±SE;n=3。肽濃度是5μM。圖45富含精氨酸的CPP誘導的神經元細胞表面結構的細胞內吞作用的建議模型的示意圖。注︰模型適用於神經元突觸和外突觸質膜和潛在的星形膠質細胞、周細胞、腦內皮細胞、少突膠質細胞和小膠質細胞的質膜。NMDA︰N-甲基-D-天冬氨酸受體;AMPAR:α-氨基-3-羥基-5-甲基-4-異惡唑丙酸受體;NCX︰鈉鈣交換體;VGCC︰電壓門控性鈣通道(例如CaV2.2、CaV3.3);ASIC︰酸敏感離子通道;TRPM2/7:瞬時受體電位陽離子通道2和7:mGluR︰代謝型穀氨酸受體;VR1︰香草酸受體1或瞬時受體電位陽離子通道亞科V成員1;TNFR︰腫瘤壞死因子受體;EAAT︰興奮性胺基酸轉運體;AQP4:水通道蛋白4;Trk:原肌球蛋白-受體-激酶受體。圖46和47是本說明書中所描述和使用的序列的參考表。具體實施方式在本說明書全文中,除非上下文另有所指,詞語「包括(comprise)」或其它變化形式,如「包括(comprises)」或「包括(comprising)」或「包含(include)」或「包含(including)」,將被理解為暗示含有陳述的整體或整體的組,但不是排除任何其他整體或整體的組。本發明涉及分離的肽和包含分離的肽的組合物,及它們的用途。所述分離的肽其特徵在於,在神經損傷或者缺血性事件前或後施用所述分離的肽可以減少神經損傷或腦血管缺血性事件(例如以及特別地,中風)的神經退行性效應。因此,施用本發明的組合物降低神經損傷或腦血管缺血性事件之後神經元細胞的損失。令人驚訝的是,申請人現在已經發現,某些CPPs形式的多肽以及具有連續鹼性/陽離子胺基酸片段(特別是精氨酸殘基,但也包括賴氨酸和色氨酸殘基)的多肽表現出神經活性或神經保護活性,並且其本身可作為神經保護劑(即用於治療神經損傷),即無需與其他神經保護劑或肽融合。因此,本發明涉及長度為10至32個胺基酸的多肽,其中10至22個胺基酸是陽離子胺基酸殘基(通常是精氨酸殘基),這種多肽通常具有精氨酸,其含量為30%或更高,這種肽在已建立的神經損傷模型中表現出神經保護活性。這包括聚精氨酸肽、硫酸魚精蛋白(如從鮭魚精子得到的魚精蛋白肽的混合物),及其這些肽的各種版本、類似物、變體或片段,包括魚精蛋白和低分子量魚精蛋白(LMWP)及其混合物。本文中使用的術語「胺基酸」或「殘基」包括二十種天然存在的胺基酸中的任意一種、任意一種天然存在的胺基酸、非天然存在的胺基酸及其衍生物、類似物和模擬物的D型。任何胺基酸,包括天然存在的胺基酸,可以是市售的或通過本領域中已知的方法合成的。非天然存在的胺基酸的例子包括正亮氨酸(「Nle」)、正纈氨酸(「Nva」)、β-丙氨酸、L-或D-萘基丙氨酸(naphthalanine)、鳥氨酸(「Orn」)、高精氨酸(homoArg)和其他肽領域公知的其他非天然存在的胺基酸,包括那些描述在M.Bodanzsky的「PrinciplesofPeptideSynthesis,」1stand2ndreviseded.,Springer-Verlag,NewYork,N.Y.,1984and1993(《肽合成原理》,第一和第二修訂版,施普林格,紐約市,紐約州,1984和1993)以及Stewart和Young的「SolidPhasePeptideSynthesis,」2nded.,PierceChemicalCo.,Rockford,Ill.,1984(《固相肽合成》,第二版,皮爾斯化學公司,羅克福德,伊利諾州,1984),二者通過引用併入本文。常見的胺基酸可以通過它們的全名,標準的單字母表示法(IUPAC),或者標準的三字母表示法表示,例如:A、Ala、丙氨酸;C、Cys、半胱氨酸;D、Asp、天冬氨酸;E、Glu、穀氨酸;F、Phe、苯丙氨酸;G、Gly、甘氨酸;H、His、組氨酸;I、Ile、異亮氨酸;K、Lys、賴氨酸;L、Leu、亮氨酸;M、Met、蛋氨酸;N、Asn、天門冬醯胺;P、Pro、脯氨酸;Q、Gln、穀氨醯胺;R、Arg、精氨酸;S、Ser,絲氨酸;T、Thr,蘇氨酸;V、Val、纈氨酸;W、Trp、色氨酸;X、Hyp、羥脯氨酸;Y、Tyr、酪氨酸。本文的組合物中任何和所有的胺基酸可以是天然存在的、合成的和其衍生物或模擬物。如本文所用到的「分離的」是指本文所描述的肽不是處於自然狀態(例如,它是從天然存在或通常相關的較大的蛋白質分子或細胞碎片中分離),或者是天然存在的蛋白質的非天然存在的片段(例如,所述肽包含少於25%,優選小於10%和更優選小於5%的天然存在的蛋白質)。分離的還可以表示肽的胺基酸序列不是自然生成的,例如,因為該序列是從天然存在的序列改性的(例如通過改變某些胺基酸,包括鹼性(即陽離子)胺基酸,例如精氨酸、色氨酸或賴氨酸,或者是因為該序列不包含天然存在的側翼胺基酸。術語「分離的」可以表示所述肽或胺基酸序列是人造序列或多肽,並且可以是非天然存在的。同樣地,「分離的」用於連接編碼上述所有肽的核酸,例如,分離的核酸是從與它們本質上相關的鄰近的核苷酸中分離,並且可以重組、合成產生,通過純化生物提取物等產生。分離的核酸可以包含編碼上述肽中的一種的一部分以及編碼另一種肽或蛋白質的另一部分。分離的核酸也可被標記。所述核酸包含優選用於動物、細菌、植物或真菌用途的密碼子。在某些實施例中,分離的核酸是載體,如表達載體,其包括編碼上述分離的肽中的一種的核酸。在,例如,BrownJ.等,(1979),MethodsinEnzymology(酶學方法),68:109;SambrookJ,ManiatisT(1989)(見上文)中提供了一種用於構建任何所需的DNA序列的通用方法。也考慮了肽的非肽類似物,例如,那些提供穩定結構或減少生物降解的肽的非肽類似物。肽模擬類似物可以根據選定的肽用非肽部分替換一個或多個殘基來製備。優選地,所述非肽部分允許肽保留它的天然結構,或穩定優選的(例如)生物活性結構。一種從肽製備成非肽模擬類似物的方法公開在Nachman等,Regul.Pept.57:359-370,(1995)。本文所用的術語「肽」包括上述所有肽。如上所述,本發明的肽可以由天然存在的胺基酸(即L-胺基酸組成)或由D-胺基酸組成(即胺基酸序列包括相對於天然序列的逆向-反向(retro-inverso)順序的D-胺基酸)。術語「逆向-反向」(「retro-inverso」)是指線性肽的同分異構體,其中序列的方向是反轉的並且每個胺基酸殘基的手性是顛倒的。因此,本文的任何序列,以L-型存在的本質上也作為D-對映體(逆向-反向)肽序列公開了。根據本發明的D-對映體(逆向-反向)肽序列是可以構建的,例如,通過合成相應的天然L-胺基酸序列的反向胺基酸序列。在D-逆向-反向對映體肽中,例如,分離的肽的一組分,交換每個單醯胺鍵的羰基和氨基的位置,而保留每一個α碳的側鏈基團的位置。製備上述限定的具有D-對映體胺基酸的本發明的分離的肽組分,可以通過化學合成相應的天然存在的L-型胺基酸序列的逆向胺基酸序列或由本領域技術人員公知的任何其它合適的方法得到。或者,肽或其組分的D-逆向-反向對映體形式可以通過使用上述利用肽或其組分的L-型作為化學合成D-逆向-反向對映體形式的基質的化學合成方法製備。富含陽離子胺基酸的多肽(其也可以稱為陽離子胺基酸的聚合物或共聚物)可以包括長度為10至32個胺基酸的多肽或低聚物。這樣,「富含陽離子」是指任何肽,寡肽,或多肽,它們通常情況下包含或包括超過30%,超過50%或甚至超過60%的陽離子殘基。在某些實施例中,可能需要多肽包括90%陽離子殘基,或甚至100%,例如,優選地精氨酸殘基。因此,富含精氨酸的多肽(其也可以稱為精氨酸的聚合物或共聚物)可以包括長度為10至32個胺基酸的多肽或低聚物。同樣地,「富含精氨酸」是指任何肽、寡肽或多肽,其包含或包括10個或更多的精氨酸殘基,或超過30%,超過50%或甚至超過60%的精氨酸殘基。因此,某些實施例包含多肽,其中100%的胺基酸是精氨酸殘基,當在R10至R18和藥學上有效的劑量的範圍內使用時具有適當的功效和較低的毒性,而在其他情況下,指其多肽具有精氨酸殘基的間歇片段(例如包括魚精蛋白和LMWP的CPPs)。通常,精氨酸殘基的片段包括連續的/相連的4至5個精氨酸殘基,被其他的胺基酸殘基穿插。在優選的實施例中,穿插的胺基酸是賴氨酸(K)殘基,因為這些殘基通常也具有陽離子電荷。在某些實施例中,精氨酸的聚合物或共聚物包括至少11個連續的精氨酸殘基,更優選至少12個連續的精氨酸殘基,更優選至少13個連續的精氨酸殘基,更優選至少14個連續的精氨酸殘基,更優選至少15個連續的精氨酸殘基,更優選至少16個連續的精氨酸殘基,更優選至少17個連續的精氨酸殘基和更優選至少18個連續的精氨酸殘基。然而,在某些實施例中,4至5個精氨酸殘基的連續序列有可能被其他的非精氨酸殘基穿插,例如本文以魚精蛋白、LMWP和它們的具有神經保護活性或用於治療神經損傷的功能性變體作為舉例的那些。在優選的實施例中,使用的是R15。連續的精氨酸殘基可以在多肽的C-端,多肽的N-端,多肽的中心(例如,被非-精氨酸胺基酸殘基包圍),或在多肽中的任意位置。非-精氨酸殘基優選不顯著降低聚合物進入細胞的膜轉運速率的胺基酸、胺基酸衍生物或胺基酸模擬物,包括(例如)甘氨酸、丙氨酸、半胱氨酸、纈氨酸、亮氨酸、異亮氨酸、蛋氨酸、絲氨酸、蘇氨酸、α-氨基-β-胍基丙酸、α-氨基-γ-胍基丁酸和α-氨基-ε-胍基己酸鹽酸。可以進行各種各樣的改變,包括以不影響肽功能的方式或有利於肽功能的方式添加不同的側基。這種改變可能涉及添加或減去電荷組,取代胺基酸,添加親油基團,其不會影響結合但是會影響促進跨過血腦屏障轉移的分子的整體電荷特性,等。對於每種改變,僅需要常規實驗以測試分子是否如本發明所述起作用。人們能簡單地作出所需的改變或選擇所需的肽,並按照實施例中詳細描述地方式應用。例如,如果肽(改性的或未改性的)在對抗紅藻氨酸的保護測試中是活躍的,或者,如果這種肽在神經遞質功能的測試中與母神經遞質競爭,則該肽是一種功能性神經遞質肽。本發明還包括所述分離的肽的功能性變體。如本文所用,分離的肽的「功能性變體」或「變體」是一種肽,其包含對分離的肽的原始胺基酸序列的一種或多種改性,並保留本文公開的性質。可作出產生分離的肽的功能性變體的改性,例如,1)加強分離的肽的性能,如在表達系統中的肽穩定性;2)為分離的肽提供新的活性或性質,如加入抗原表位或加入可檢測部分;或3)提供產生相同或相似的肽性質的不同胺基酸序列。對分離的肽改性可以製成編碼肽的核酸,並且可以包括刪除、點突變、截短、胺基酸取代和胺基酸添加。或者,可以直接對肽改性,例如通過切割、添加連接分子(優選可切割的接頭,例如MMP、鈣蛋白酶、tPA)、添加可檢測的部分(如生物素)、添加脂肪酸、一種胺基酸取代為另一種,等等。改性還包括融合蛋白,其包括全部或一部分所述分離的肽的胺基酸序列。在一個實施例中,接頭選自一個或多個MMP型接頭、鈣蛋白酶、半胱天冬酶或tPA接頭。MMP型接頭定義為由基質金屬蛋白酶(MMPs)識別和裂解的肽序列。同樣地,鈣蛋白酶、半胱天冬酶或tPA接頭是由這些蛋白酶識別和裂解的肽序列。在某些實施例中,本發明的多肽還融合到其他多肽,這些其他多肽被轉運到神經損傷的位點或在神經細胞內轉運。同樣的,本發明的多肽也可以連接到輔助肽,該輔助肽可以與對神經功能有害的酶結合或者相互作用,所以所述輔助肽可以作為以下通過本發明的肽跨過細胞膜轉運的酶的競爭性抑制劑。輔助肽是tPA、鈣蛋白酶和MMP,和使用可裂解的接頭(如半胱天冬酶序列)連接到本發明的肽,因此tPA、鈣蛋白酶或MMP從本發明的肽中釋放出來,然後作為對神經功能有害的酶的細胞內競爭性抑制劑。同樣的,本發明延伸到一種本發明的肽,如R10-R18或Ptm1-5或LMWP或它們的變體,連接於半胱天冬酶裂解位點,其本身又與鈣蛋白酶、tPA或MMP連接。本文所定義的術語「序列同一性」意味著序列進行如下比較。為了確定兩個胺基酸序列的同一性百分比,為了最佳比較目的對序列比對(例如,可以在第一胺基酸序列的序列中引入間隙)。然後比較在相應的胺基酸位置的胺基酸。當第一序列中的一個位置被第二序列中相應位置的相同胺基酸佔據時,則該分子在該位置是相同的。兩個序列之間的同一性的百分比是序列共有的相同位置的數目的函數。例如,據說一個特定的肽相對於具有明確長度的參比多肽具有具體的同一性百分比,所述同一性百分比是相對於參比肽。因而,一種肽與長度為100個胺基酸的參比多肽有50%相同,可以是多肽的50個胺基酸與參比多肽的長度為50個胺基酸的部分完全相同。它也可能是一個長度為100個胺基酸的多肽,它的整個長度有50%與參比多肽相同。這樣兩個序列的同一性百分比的測定可以使用數學算法來完成。優選的,一種用於比較兩序列的數學算法的非限制性例子是Karlin等的算法,(1993),PNASUSA,90:5873-5877。這樣的算法併入NBLAST程序,可以用來確定與本發明的胺基酸序列具有所需要的同一性的序列。為了比較目的得到缺口對齊,可以使用缺口BLAST如Altschul等,(1997),NucleicAcidsRes,25:3389-3402中所描述的。當使用BLAST和缺口BLAST程序時,可使用各自程序(例如,NBLAST)的預設參數。該序列可以利用GeneticComputingGroup's(遺傳計算組)的GAP(globalalignmentprogram(全局比準程序))第9版進一步對齊,使用預設(BLOSUM62)矩陣(值-4至+11)與-12的缺口開放罰分(對缺口的第一空缺)和-4的缺口延伸罰分(在缺口中每一個額外的連續的空缺)。對齊後,同一性百分比是通過表達匹配的數目作為所要求保護的序列中的胺基酸數目的百分率來計算。所描述的測定兩個胺基酸序列的同一性百分比的方法可以相應地適用於核酸序列。本發明的肽可以直接連接或通過接頭連接。在本上下文中的「接頭」通常是肽、寡肽或多肽,並可以用於將多個多肽彼此連接。選擇彼此連接的本發明的多肽可以是相同的序列,或選自本發明的任何多肽。接頭可以有1-10個胺基酸的長度,更優選1至5個胺基酸的長度,最優選1至3個胺基酸的長度。在某些實施例中,接頭不需要任何有二級結構形成性能,即不需要α螺旋或β片狀結構形成傾向,例如,接頭是由至少35%的甘氨酸殘基組成。正如上文所述的,接頭可以是可裂解的肽,例如MMP肽通過正常細胞過程可以在細胞內裂解,有效提高預先連接肽的細胞內的劑量,同時保持細胞外劑量足夠低,低至不被認為是有毒的。使用一種細胞內/內源性可裂解的肽、寡肽或多肽序列作為接頭,允許多肽在遞送到目標細胞後彼此分離。在這種情況下可裂解的寡肽或多肽序列還包括蛋白酶可裂解的寡肽或多肽序列,其中通常選擇蛋白酶裂解位點是依賴於經處理的細胞內源性表達蛋白酶。如上定義的接頭,如果以低聚或多肽序列存在,可以由D-胺基酸或天然存在的胺基酸(即L-胺基酸)組成。作為上述的替代物,多肽的偶聯或融合可以通過耦合劑或連接劑(例如交聯劑)來完成。可以使用幾種分子間的交聯劑,例如,Means和Feeney,ChemicalModificationofProteins(蛋白質的化學改性),Holden-Day,1974,第39-43頁。這些試劑是,例如,N-琥珀醯亞胺-3-(2-吡啶基二硫)丙酸酯(SPDP)或N,N'-(1,3-亞苯基)雙馬來醯亞胺;N,N'-乙烯-雙-(碘乙醯胺)或其他這種具有6至11個碳亞甲基橋的試劑;和1,5-二氟-2,4-二硝基苯。用於此目的其他試劑包括:p,p'-二氟-m,m'-二硝基聯苯碸;二甲基己二亞醯胺化物;酚-1,4-二磺醯氯;六亞甲基二異氰酸酯或二異硫氰酸酯,或偶氮苯基-p-二異氰酸酯;戊二醛和二重氮聯苯胺。交聯試劑可以是同型雙功能,即具有進行相同的反應的兩個官能團。優選的同型雙功能交聯是二馬來醯亞胺己烷(BMH)。BMH含有兩個馬來醯亞胺官能團,其在溫和條件下(pH值6.5-7.7)與含巰基的化合物特異性地反應。兩個馬來醯亞胺基團是通過烴鏈連接的。因此,BMH對包含半胱氨酸殘基的蛋白質(或多肽)的不可逆交聯有用。交聯劑也可以是異型雙功能。異型雙功能交聯劑有兩個不同的官能團,例如胺反應基和硫醇反應基,將分別與具有游離胺和硫醇的兩種蛋白質交聯。異型雙功能交聯劑的例子是琥珀醯亞胺基4-(N-馬來醯亞胺甲基)環己烷-1-羧酸酯(SMCC)、m-馬來醯亞胺苯甲醯基-N-羥基琥珀醯亞胺酯(MBS)和琥珀醯亞胺4-(p-馬來醯亞胺苯基)丁酸酯(SMPB、MBS的擴鏈類似物。這些交聯試劑的琥珀醯亞胺基具有伯胺以及硫醇反應的馬來醯亞胺,與半胱氨酸殘基的硫醇形成共價鍵。因為交聯試劑通常在水中的溶解度低,將親水性部分如磺酸鹽基團添加到交聯劑中,以提高其水溶性。硫代MBS和硫代SMCC是交聯劑水溶性改性的例子。許多交聯試劑產生共軛,在細胞條件下基本上是不可裂解的。因此,一些交聯試劑含有共價鍵,如二硫鍵,其在細胞條件下可裂解。例如,Traut’s試劑、二硫基(丙酸琥珀醯亞胺)(DSP)和N-琥珀醯亞胺3-(2-吡啶基二硫)丙酸(SPDP)是公知可裂解的交聯劑。可裂解的交聯試劑的使用允許多肽在遞送到靶細胞後分離,必要時,提供的細胞能夠裂解交聯劑的特定序列。為此目的,直接的二硫鍵聯接也可能是有用的。化學交聯還可包括使用間隔臂。間隔臂提供了分子內的靈活性或調整共軛基團之間的分子內距離,從而可能有助於保護生物活性。間隔臂可以以包含間隔胺基酸的蛋白質(或多肽)的一部分的形式,例如:脯氨酸。或者,間隔臂可能是交聯劑的一部分,如在「長鏈SPDP」(PierceChem.Co.,Rockford,Ill.,cat.No.21651H)。許多交聯劑,包括上面討論的,是市售的。它們使用的詳細說明很容易從商業供應商獲得。蛋白質交聯和共軛製備的一般參照:Wong,ChemistryofProteinConjugationandCross-Linking,CRCPress(1991)。本發明的多肽也可以包含「衍生物」、「變體」或「功能性片段」,即一種肽的序列來自上文定義本發明的肽的天然存在(L-胺基酸)的序列,作為胺基酸序列的一個或多個位點上的一個或多個胺基酸的取代,作為在天然存在序列的任何位點上的一個或多個胺基酸的刪除,和/或作為在天然存在的肽的序列的一個或多個位點上的一個或多個胺基酸的插入。如果用作本發明的多肽,「衍生物」應保留其生物活性,例如,任何本發明的多肽衍生物,應保留其神經保護活性。在本發明的上下文中的衍生物也可以以上文限定的它們的L-或D-胺基酸序列的形式存在,或兩者共存。如果胺基酸的取代是用於製備本發明的多肽的衍生物,優選保守(胺基酸)取代。保守(胺基酸)取代通常包括以下組中的取代︰甘氨酸和丙氨酸;纈氨酸、異亮氨酸、亮氨酸;天冬氨酸和穀氨酸;天冬醯胺和穀氨醯胺;絲氨酸和蘇氨酸;賴氨酸和精氨酸;苯丙氨酸和酪氨酸。因此,優選的保守取代組是天冬氨酸-穀氨酸;天冬醯胺-穀氨醯胺;纈氨酸-亮氨酸-異亮氨酸;丙氨酸-纈氨酸;和苯丙氨酸-酪氨酸。通過這種突變例如肽的穩定性和/或有效性可能會提高。如果肽中引入突變,肽保持(功能上)一致,例如在序列上,在功能上,和在抗原特性上或其他功能。這種肽的突變部分可以擁有改變了的性能,相對本發明的多肽的非改變序列在某些應用上(例如增加最優pH值、增加溫度穩定性等)可能更具優勢。本發明的肽的衍生物被認為與本發明的肽的未改性序列具有實質同一性。特別優選的胺基酸序列相對於天然存在的類似物具有至少30%序列同一性,優選至少50%序列同一性,甚至優選至少60%序列同一性,甚至優選至少75%序列同一性,甚至更優選至少80%同一性,但更優選90%同一性和更優選至少95%或甚至99%序列同一性。上文描述了本發明的多肽的功能衍生物的合成或分離的適當方法,以及用於確定兩個胺基酸序列的同一性百分比的方法。此外,上述公開的肽衍生物的生產方法是公知的,以及可以根據以下本領域技術人員公知的標準方法進行(例如,參見SambrookJ,ManiatisT(1989))。作為進一步的實施例,本發明提供了包含本文所限定的肽的藥物組合物或藥物。在某些實施例中,這種藥物組合物或藥物包含本文所限定的肽以及任意的接頭。此外,這種藥物組合物或藥物可以包括藥學上可接受的載體、助劑或媒介物。根據本發明「藥學上可接受的載體、助劑或媒介物」指無毒的載體、助劑或媒介物,配製時不會破壞肽的藥理活性或生理靶向性。可用於本發明的藥物組合物的藥學上可接受的載體、助劑或媒介物包括但不限於那些可用於頭顱或頭顱內,或可以穿過血腦屏障(BBB)的載體、助劑或媒介物。不過,本發明的藥物組合物可以包括,離子交換劑、氧化鋁、硬脂酸鋁、卵磷脂、血清蛋白(如人血清白蛋白)、緩衝物質(如磷酸鹽)、甘氨酸、山梨酸、山梨酸鉀、飽和植物油脂肪酸的部分甘油酯混合物、水、鹽或電解質(如磷酸氫二鈉、磷酸氫鉀、氯化鈉、鋅鹽)、矽膠、三矽酸鎂、聚乙烯吡咯烷酮、纖維素基物質、聚乙二醇、羧甲基纖維素鈉、聚丙烯酸酯、蠟、聚乙烯-聚氧丙烯嵌段聚合物、聚乙二醇和羊毛脂。本發明組合物可口服給藥、腸道外給藥、通過霧化吸入、局部給藥、直腸給藥、鼻腔給藥、口腔給藥、陰道給藥、腦給藥或通過植入儲庫給藥。本文中的術語「腸道外」包括皮下、靜脈、肌肉、關節內、滑膜內、胸骨內、鞘內、肝內、病灶內以及顱內注射或輸注技術。該藥物組合物通過口服給藥、腹膜內給藥或靜脈給藥。本發明的藥物組合物的無菌可注射形式可以是水性或油性懸浮液。這些懸浮劑可根據本領域公知技術,使用適當的分散劑或溼潤劑和懸浮劑進行配製。無菌可注射的製劑還可以是在無毒的腸道外給藥可接受的稀釋液或溶劑中的無菌可注射劑溶液或懸浮液,例如,在1,3-丁二醇中的溶液。可接受的媒介物和溶劑可以是水、Ringer's溶液和等滲氯化鈉溶液。此外,無菌的固定油通常作為溶劑或懸浮介質。同樣地,可以使用任何溫和的不揮髮油,包括合成的單或雙甘油酯。脂肪酸例如油酸及它的甘油酯衍生物在可注射製劑的配製中是有用的,因為它們是天然的藥學上可接受的油,如是橄欖油或篦麻油,尤其以它們的聚氧乙烯化的形式。這些油溶液或懸浮液還可以包含長鏈醇稀釋劑或分散劑,例如羧甲基纖維素或類似的分散劑,其通常用於藥學上可接受的劑型,包括乳劑和混懸劑製劑。另外常用的表面活性劑,例如吐溫、司盤和其他乳化劑或生物利用度增強劑,它們通常用於如常用於製造藥學上可接受的固體、液體或也可用於本發明的配製其他劑型。本文中藥學上可接受的組合物可以以任何口服可接受的劑型通過口服給藥,包括但不限於,膠囊、片劑、水性懸液或溶液。口服使用片劑的情況下,常用的載體包括乳糖和玉米澱粉。通常還加入潤滑劑,如硬脂酸鎂。以膠囊形式口服給藥,有用的稀釋劑包括乳糖和乾燥的玉米澱粉。當需要口服水性懸液時,活性成分與乳化劑和懸浮劑結合。必要時,還可加入某些甜味劑、調味劑或著色劑。或者,本文所定義的藥物組合物可以栓劑的形式用於直腸給藥。這種栓劑可以通過藥物與適當的的無刺激性的輔料混合配製,該輔料在室溫下是固體但在直腸溫度下為液體,因此,將在直腸中融化以釋放藥物。這類材料包括,可可油、蜂蠟和聚乙二醇。如本文所定義的藥物組合物也可以通過局部給藥,尤其當治療目標包括區域或器官更容易通過局部施用,包括腦、其它顱內組織、眼或皮膚疾病。能夠容易地製備適當的製劑用於這些區域或器官中的每個。對於局部施用,本文所定義的藥物組合物可製備成合適的藥膏,所述藥膏包含本文所限定的肽、懸浮或溶解在一種或多種載體中。肽的局部給藥的載體包括但不限於,礦物油、液態石蠟、白凡士林、丙二醇、聚氧乙烯、聚氧丙烯化合物、乳化蠟和水。或者,本文限定的藥物組合物可製備成合適的洗劑或面霜,其包含懸浮或溶解在一種或多種藥學上可接受的載體中的肽。合適的載體包括但不限於,礦物油、山梨醇酐單硬脂酸酯、聚山梨醇酯60、十六醇酯蠟、棕櫚醇、2-辛基十二醇、苄醇和水。本文限定的的藥物組合物還可通過鼻腔氣霧劑或吸入劑給藥。這種組合物可以根據藥物劑型領域中公知的技術配製,並且可配製成在生理鹽水中的溶液,採用苄醇或其他合適的防腐劑、提高生物利用度的吸收促進劑、碳氟化合物和/或其他常見的增溶劑或分散劑。本文中藥學上可接受的組合物或藥物被配製用於口服或腸胃外給藥,例如通過注射。為了治療的目的,無毒、減少損傷、有效量的肽可用於製備上述限定的藥物組合物。因此,一些肽可以與載體材料結合以製備如上所限定的組合物。該藥物組合物通常製備成單(或多)劑量的形式,這將根據所治療的宿主和給藥的特定模式而變化。通常,藥物組合物按下述配製,使施用給接受藥物組合物的患者的劑量範圍為每劑量0.0001至100毫克/公斤體重/天的肽。優選的劑量範圍為每劑量0.01毫克/公斤體重/天至50毫克/公斤體重/天,甚至進一步優選的劑量範圍為每劑量從0.1毫克/公斤體重/天至10毫克/公斤體重/天。然而,上述的劑量範圍和治療方案可以適宜於任何特定患者,取決於各種因素包括使用具體的肽的活性、年齡、體重、總體健康狀況、性別、飲食、給藥時間、排洩率、藥物組合、治療醫師的診斷和被治療的特定疾病的嚴重程度。在這種情況下,可以在初始劑量範圍內給藥,劑量可以隨著治療時間而變化,例如,在如前所述的範圍內增加或減少初始劑量範圍。或者,可以通過施用具體的劑量範圍以連續的方式給藥,從而在整個治療時間保持初始劑量範圍。二者的給藥形式可進一步組合,例如,假如劑量範圍適應(增加或減少)治療的不同時期,但單個時期內保持恆定,以便不同時期的劑量範圍彼此不同。本發明的藥物組合物和/或肽可以用於治療、改善或預防與損傷細胞的損傷效應相關的疾病,特別是哺乳動物細胞,如本文所公開的,尤其是用於神經損傷的治療,包括腦中風或脊髓損傷、癲癇、圍產期缺氧缺血、腦或脊髓的缺血性或創傷性損傷和中樞神經系統(CNS)中的神經元損害包括但不限於,急性CNS損傷、腦缺血性中風或脊髓脊髓損傷以及缺氧、缺血、機械損傷、神經性疼痛、興奮性毒性以及相關的損傷。此外,本發明的藥物組合物和多肽可用於提供神經活性或神經保護,對抗或者治療興奮性毒性和缺血性損傷、興奮性中毒、缺乏神經營養支持、神經元斷裂、神經元損傷,包括例如癲癇、慢性神經退行性病症和類似疾病。在上下文中,興奮性毒性可以尤其涉及中風、創傷性腦損傷和中樞神經系統(CNS)的神經變性疾病,如多發性硬化(MS)、阿爾茨海默氏病(AD)、肌萎縮性側索硬化症(ALS)、神經性疼痛、纖維肌痛、帕金森氏病(PD)、圍產期缺氧缺血和亨廷頓氏病,本發明均可以治療。引起神經元周圍過多的穀氨酸濃度以及本發明可治療的其他常見症狀是低血糖、腦血管痙攣、苯並二氮雜戒斷和癲癇持續狀態、視網膜神經節細胞的青光眼/退化,等等。以上限定的與損傷哺乳動物細胞的損傷效應相關的疾病以及本文中提到的進一步的疾病或者紊亂的治療、改善或預防,通常是在本發明所描述的劑量範圍內施用本發明的藥物組合物或肽或肽混合物。所述藥物組合物或肽可以在興奮性中毒和/或(缺血性)腦損傷即損傷哺乳動物細胞的損傷效應發作之前施用,或其同時或隨後進行;例如,施用藥物組合物或肽可以在腦中風或脊髓損傷、腦或脊髓的缺血性或創傷性損傷,通常,中樞神經系統(CNS)神經元損傷後的(長達)1小時(0-1小時)、長達2小時、長達3-5小時或長達24小時或更長時間內進行。在慢性神經退行性疾病(AD、PD、ALS、MS等)治療可能需要終身每日治療。本發明的化合物用於治療時,施用治療有效量。通常,治療有效量是指延遲疾病發作,抑制疾病進展,或治癒所治療的全部特定症狀所需要的劑量。治療有效量具體是指那些理想地能影響中風或其他腦缺血性損傷後的神經元的存活的劑量。通常,治療有效量將隨著受試者的年齡和身體狀況以及受試者疾病的性質和程度而變化,所有這些可以由本領域的普通技術人員決定。劑量可由個體醫生進行調整,特別是在正在經歷任何併發症的情況下。如上所述,本發明的一個方面涉及編碼分離的肽或其變體的核酸序列以及它們的衍生物,以及在嚴格條件下與由上述描述的核苷酸序列組成的核酸分子雜交的其他核苷酸序列。本文所用的術語「嚴格條件」,是指與本領域所熟悉的參數。核酸雜交參數在彙編這些方法的參考文獻中可以找到,例如:MolecularCloning:ALaboratoryManual,J.Sambrook,etal.,eds.,SecondEdition,ColdSpringHarborLaboratoryPress,ColdSpringHarbor,N.Y.,1989,(分子克隆:實驗指南,J.Sambrook等,第二版,冷泉港實驗室出版社,冷泉港,紐約,1989年)或者CurrentProtocolsinMolecularBiology,F.M.Ausubel,etal.,eds.,JohnWiley&Sons,Inc.,NewYork(分子生物學現代方法,F.M.Ausubel等,JohnWiley&Sons公司,紐約)。更具體的說,本發明中所用到的嚴格條件,是指在65℃下,在雜交緩衝液(3.5×SSC,0.02%聚蔗糖,0.02%聚乙烯吡咯烷酮,0.02%牛血清白蛋白,25mMNaH2PO4(pH7),0.5%SDS,2mMEDTA)中雜交。SSC是0.15M氯化鈉/0.15M檸檬酸鈉,PH=7;SDS是十二烷基硫酸鈉;EDTA是乙二胺四乙酸。雜交後,轉移了DNA的膜在室溫下用2×SSC清洗,然後在65℃下用0.1×SSC/0.1×SDS清洗。此外本發明還提供了試劑盒,其包栝至少一種以上所述劑型中的上述藥物組合物(在一個或多個容器)和關於用法說明的使用說明書或信息手冊和/或關於藥物組合物應用的信息。在本說明書中,除非特別說明或者上下文另有規定,提到單步驟、物質組分、步驟的組或物質組分的組應包括那些步驟、物質組分、步驟的組或物質組分的組中的一個和數個(即一個或多個)。本領域的的技術人員將理解本文所述的本發明易受變體和改性的影響,不同於其他領域所描述的。需要了解的是本發明包括所有這些功能的變化和改性。本發明也包括本說明書中分別或共同地涉及或指出的所有步驟、特徵、組合物和化合物,以及任何和所有的組合或任何兩個或兩個以上的所述的步驟或特徵。本發明並不受本文中具體實施例所描述的範圍的限制,具體實施例只是為了例證。如本文所述,等效功能的產品,組合物和方法顯然是在本發明的範圍內。而且,除非另有指示,本發明不需要過度的使用分子生物學、微生物學、神經生物學、病毒學、DNA重組技術、溶液中的肽合成、固相肽合成、免疫學的常規技術,或者本文所引用的技術進行實驗。出乎意料的是,申請人發現某些CPPs,尤其是富含精氨酸的肽顯示出神經保護功效,而不需要常規地將其融合到預先鑑定的神經保護肽。這些CPPs中的某些也顯示出低毒性並且在低劑量或低濃度是有效的。這些CPPs包括穿膜肽和Pep-1。然而,更出乎意料的是,申請人還發現長鏈型鹼性(如陽離子)胺基酸,例如長度為10至20個(含)殘基的聚精氨酸肽,優選長度為10至18個(含)殘基的聚精氨酸肽,和某些包含超過10個精氨酸殘基的肽,例如魚精蛋白或LMWP,與這些CPPs相比時,以及尤其與相似濃度的較短的富含精氨酸序列(如R1到R8)相比,顯示出能極大地增強神經保護活性。特別是,與上述的CPPs相比,在穀氨基酸以及或體外局部缺血損傷模型中試驗時,R12、R15、R18均可提高神經保護功效。這兩種不同的神經元損傷模型可能激活不同的損傷細胞通路,因此將進一步了解到神經保護範圍和本發明的肽的可能作用方式。在本說明書中,縮寫詞「Arg」後的整數表示肽中精氨酸重複序列的數量。因此,Arg-15(縮寫為R15,根據IUPAC命名是精氨酸單字母縮寫)指在一個肽結構中連續的精氨酸殘基。然而,只要是可行的,本說明將採用單字母胺基酸代碼表示,即,R15,例如用於代替Arg-15。涉及神經元細胞死亡的神經障礙神經障礙,比如偏頭疼、中風、外傷性腦損傷、脊椎損傷癲癇、圍產期低氧-局部缺血和神經退行性疾病,包括亨廷頓氏病(HD)、帕金森氏病(PD)、阿爾茨滋海默氏病(AD)和肌萎縮性脊髓側索硬化症(ALS),是由長期的腦損傷引起的發病和傷殘的主要原因。通常來說,腦損傷涉及一系列的細胞死亡過程,包括細胞凋亡、自噬、壞死性凋亡和細胞壞死,以及影響神經元星形膠質細胞、少突細胞、小膠質細胞和血管內皮細胞(統一被稱為神經與血管單元;NVU)。涉及神經損傷的破壞性的觸發涉及多種途徑,包括穀氨酸鹽興奮中毒、鈣過量、氧化應激、蛋白水解酶和線粒體失調。本文所使用的術語「中風」包括任何影響腦或脊髓的缺血性紊亂,如腦或脊髓動脈中血栓栓塞性閉塞、嚴重的低血壓、圍產期低氧-局部貧血、心肌梗死、低氧、腦出血、血管痙攣、周圍血管病症、靜脈血栓、肺栓、短暫性腦缺血發作、肺部缺血、不穩定心絞痛、可逆性缺血性神經損傷、附屬溶栓作用、過度凝血條件、腦再灌注損傷、鐮狀細胞性貧血、中風障礙或醫源性引發的缺血期,如血管成形術或腦部缺血。增加神經遞質穀氨酸鹽的細胞外水平經由興奮中毒引起的急性和延遲破壞性過程可引起神經元細胞死亡。細胞外穀氨酸鹽的積累過度刺激NMDA和AMPA受體,以及隨後的VGCCs、NCX、TRMP2/7、ASIC和mGlu受體導致細胞體外的鈣離子和鈉離子內流,並從細胞儲存庫中釋放結合鈣。NMDA受體的過度激活也會觸發破壞性分子(如氧化氮、CLCA1;活化鈣氯離子通道調節器1、鈣蛋白酶,SREBP1:結合蛋白質-1的膽固醇調節器)的生成以及信號通路(如:DAPK;與死亡有關的蛋白激酶,CamKII:鈣離子-鈣調蛋白-依賴的蛋白激酶II)。細胞內鈣的增加引發一系列細胞破壞性事件,包括磷脂酶、蛋白酶、碳酸酶、激酶和一氧化氮合成酶,以及觸發細胞死亡通路(如細胞凋亡、自噬、壞死性凋亡和細胞壞死)的激活。因為本文所示的本發明的多肽保護神經元避免死亡,在WO2009133247和EP1969003中公開了本發明的肽還能用於治療和/或預防阿爾茨海默氏病(AD)、帕金森氏病(PD)、肌萎縮性脊髓側索硬化、中風、外周神經病或癲癇,還有相應的本文所述的其他相關病理學。因此,本發明針對於治療帕金森氏病、阿爾茨海默氏病、肌萎縮性脊髓側索硬化症、中風、外周神經病、癲癇、脊椎損傷、糖尿病或藥物成癮的方法,其中本發明的藥學有效量的任意一種或多種本發明的肽施用給患者。換句話說,根據本發明的肽可用於治療與局部缺血相關的損傷,還有阿爾茨海默氏病(AD)、亨廷頓氏病(HD)、帕金森氏病(PD)、多發性硬化症(MS)、急性播散性腦脊髓膜炎(ADEM)和肌萎縮性脊髓側索硬化症(ALS)、中風,外周神經病、癲癇、脊椎損傷和其他本文所述的相關病理學。在藥物應用上,肽也可以包埋在微膠囊中,例如通過共堆積技術或界面聚合(例如分別為羥基甲基纖維素或明膠微膠囊和聚-(甲基丙烯酸甲酯)微膠囊),在膠體藥物釋放系統中(例如,膠質體白蛋白微球、微乳液、納米微粒和納米膠囊),或粗乳中。這類技術公開在雷明頓的藥物科學(Remington'sPharmaceuticalSciences)中。也可利用緩釋製劑完成。緩釋製劑的適當例子包括含有肽的固體疏水性聚合物的半滲透基質,該基質是以成型製品的形式,例如膜或微膠囊。緩釋基質的例子包括在Langer等的J.Biomed.Mater.Res.,15:167-277(1981)以及Langer,Chem.Tech.,12:98-105(1982)中描述的多元酯、水凝膠,或者聚乙烯醇、聚乳酸(美國專利號3,773,919、EP58,481),或不可降解的乙烯-醋酸乙烯酯。在一個實施例中,藥物組合物包括如上所述的本發明的肽,用於治療缺血性損傷、阿爾茨海默氏病、帕金森氏病、肌萎縮性脊髓側索硬化症(ALS)、中風、外周神經病或癲癇。在另一個實施例中,本發明提供了促進神經元存活的方法,該方法包括將本發明的肽或其組合物與神經元接觸的步驟。如本文所示該方法可在體外完成。本文中所用的出版物以及其他材料用於闡明本發明的背景,特別是提供與實踐相關的其他細節,均是通過引用包含在本文中。本發明在以下例子中進一步描述,但並不限制本發明的範圍。實例(下表1、2和5的聚精氨酸和富含精氨酸和富含精氨酸的魚精蛋白肽)硫酸魚精蛋白、魚精蛋白肽和其他肽表5中列出的肽,硫酸魚精蛋白(魚精蛋白;Ptm)得自SanofiAventis。小分子量魚精蛋白(LMWP)是由MimotopesPtyLtd(邁拓公司)(澳大利亞)合成的。魚精蛋白肽1-5(Ptm1、Ptm2、Ptm3、Ptm4、Ptm5)由PepmicCoLtd(中國)合成。所述肽由HPLC純化,純度大於90-98%。所有的肽製備成在生理鹽水中的100X儲存液(500μM),並且根據損傷模型在濃度範圍為0.1-10μM中進行評估。需要注意的是硫酸魚精蛋白(魚精蛋白;Ptm)是Ptm1-Ptm41的混合物。魚精蛋白肽(Ptm、Ptm1-5、LMWP)富含精氨酸。方法(聚精氨酸和富含精氨酸的肽)原代神經元皮質培養皮質培養的建立如現有技術所描述(Melon等,2001)。簡而言之,來自E18-E19Sprague-Dawley大鼠的皮質組織是在添加有1.3mML-半胱氨酸,0.9mMNaHCO3,10units/mL木瓜蛋白酶(Sigma,美國)和50units/mL脫氧核糖核酸酶(Sigma)的良伊格爾培養基(DMEM;Invitrogen,澳大利亞)中分離,用冷DMEM/10%馬血清衝洗。神經元重懸在含2%B27補充物(B27;Invitrogen)的Neurobasal(Invitrogen)(神經基礎培養基(Invitrogen))中。在接種前,96-孔型玻璃管孔(直徑6mm,ProTech,澳大利亞)或96-孔塑料板(ProSciTech,澳大利亞)表面塗覆有聚-D-賴氨酸過夜(50ml/孔:50mg/mL;70-150K,Sigma)。然後去除過量的聚-D-賴氨酸,並用Neurobasal(含2%B27;4%胎牛血清;1%馬血清;62.5mM穀氨酸鹽;25mM2-巰基乙醇;和30mg/mL鏈黴素和30mg/mL盤尼西林)替換。體外培養第11-12天,神經元鋪板,每個孔有約10000個活的神經元。神經元保持在37℃的CO2培養箱(5%CO2,95%大氣平衡,98%溼度)中,在體外培養第4天,移除1/3的培養基,並用含有絲分裂抑制劑和胞嘧啶阿拉伯呋喃糖苷(1mM終濃度;Sigma)的新鮮的Neurobasal/2%B27替換。在體外培養第8天,將一半的培養基用Neurobasal/2%B27替換。在體外培養第11或12天之後,培養物通常由>97%神經元和1-3%星形膠質細胞組成(Meloni等,2001)。使用的其他細胞系腦內皮細胞系(bEND3)和神經母細胞瘤細胞系(SH-SY5Y)也可用於某些實驗中。使用標準技術在DMEM中加入5或15%的胎牛血清培養bEND3和SH-SY5Y細胞。小鼠原代星形膠質細胞的獲得和培養,如培養皮層神經元所描述的,除了使用DMEM+10%胎牛血清代替Neurobasal/2%B27外,不使用有絲分裂抑制劑、胞嘧啶阿拉伯呋喃糖苷。細胞穿膜肽和對照肽表1中列出的肽通過MimotopesPtyLtd(邁拓公司)(澳大利亞)合成,除了TAT-L(其是由PepscanPresto(派普斯坎公司)(荷蘭)合成)以及R9/tPA/R9、NCXBP3和R9/X7/R9(它們由ChinaPeptideCo.,Ltd(中肽生化有限公司)合成)。列於表2中的肽由ChinaPeptides(中肽生化有限公司)(中國)合成。所述肽由HPLC純化到純度大於88-96%。TAT-L、穿膜肽、R9和Pep-1合成為抗蛋白酶D-逆向-反向形式的L-同種型、TAT-D和Arg9-D,是從D-胺基酸的逆次序合成的(下文中稱為D-同種型)(Brugidou等,1995)(表1)。採用D-同種型的融合TAT的JNK抑制肽(JNKI-1D-TAT)以及L-同種型的融合TAT的AP-1抑制肽(PYC36L-TAT)作為陽性對照(表1;Borsello等,2003;Meade等,2010b)。所有的肽製備成在生理鹽水中的100X儲存液(500μM),並依據損傷模型來在濃度範圍0.1-15μM進行評估。TAT-L肽僅用於穀氨酸興奮中毒模型中。表1:各肽的胺基酸序列、分子量和電荷在N-端上,H代表游離胺,和Ac代表乙醯基。在C-端上,OH代表游離酸,NH2代表醯胺。AA=胺基酸。小寫字母代碼指代胺基酸的D-同種型。a肽在Hoetal(2001)(Ho等(2001))中有描述,bNR2B9c-TAT也已知為NA-1(Aarts等,2002;Hill等,2012),c肽在Meade等,2010ab中有描述並由PhylogicalLtd進行了分離,d肽在Borsello等,2003中有描述,eXIP由He等,1997進行了描述,f肽是JaneCross/BrunoMeloni從phylomer肽庫(PhylogicaPtyLtd)分離得到。穀氨酸和紅藻氨酸和NMDA興奮性中毒模型和肽的培養在暴露於穀氨酸或紅藻氨酸前15分鐘,通過去除培養基並添加50μl含有CPPs、對照肽或MK801/CNQX的Neurobasal/2%B27,將肽加到培養孔(96-孔板式)。為誘導興奮性中毒,將50μl含有穀氨酸(200μM)或紅藻氨酸(400μM)或NMDA(200μM)的Neurobasal/2%B27加入到培養孔中(100μM穀氨酸、200μM紅藻氨酸和NMDA100μM終濃度)。培養物在37℃、CO2培養箱中進行培養,對於穀氨酸暴露來說,培養5分鐘,對於紅藻氨酸暴露來說,培養45分鐘,對於NMDA暴露來說,培養10分鐘,該段時間之後,用100μl的50%Neurobasal/2%N2補充物(Invitrogen)和50%平衡鹽溶液(BSS;見下文)替換培養基。將培養物在37℃的CO2培養箱中進一步培養24小時。採用或未採用穀氨酸或紅藻氨酸處理的未處理對照接受相同的清洗步驟並添加相同培養基。在一個實施例中,在15分鐘CPP培養(5或10μM)後,去除孔中培養基並立刻用300μlBSS清洗,然後添加含穀氨酸(100μM/100μl)的Neurobasal/2%B27。之後,按照之前所述的方法處理培養物。接受或未接受穀氨酸暴露的未處理對照組接受相同的清洗步驟並添加相同的培養基。另外,進行R9肽和JNKI-1D-TAT對照肽的穀氨酸暴露後CPP處理(5μM)實驗(post-glutamicacidexposureCPPtreatment(5μM)experiment)。在這個實驗中,如上所述,神經元暴露於包含在100μlNeurobasal/2%B27中的穀氨酸(100μM)5分鐘,該段時間之後去除培養基,並用50μlNeurobasal/2%N2補充物代替,然後,暴露於穀氨酸的第0分鐘和第15分鐘,加入肽(在BSS中10μM/50μl)。對於穀氨酸暴露前實驗(pre-glutamicacidexposureexperiment),神經元暴露於肽10分鐘,緊接在暴露於穀氨酸前或者暴露於穀氨酸前的1小時、2小時、3小時、4小時或5小時。這一步可通過去除培養物和加入50μl含肽的Neurobasal/2%B27培養基完成。在37℃的CO2培養箱中10分鐘後,去除培養基並用100μL的Neurobasal/2%B27代替(對於含穀氨酸的直接暴露於穀氨酸的培養基,100μM)。在相關肽的預處理期間,去除培養基,用100μl含穀氨酸(100μM)的Neurobasal/2%B27培養基替換。暴露於穀氨酸5分鐘後,神經元培養孔按上述方法進行處理。對於所有試驗,採用或未採用穀氨酸處理的未處理對照都經過相同的培養步驟並添加相同的培養基。肝素實驗肝素(注射)從Pfizer(1000IU/ml)獲得。進行兩種不同的肝素實驗:1.肽與肝素(20IU/ml)在Neurobasal/B27中室溫下培養5分鐘,在添加到培養孔(50μl)前,在37℃的CO2培養箱中培養15分鐘。培養後,去除孔中的培養基,並用100μl的含穀氨酸(100μM)的Neurobasal/2%B27替換,隨後的處理如上所述;2.孔中的培養基用含肝素(50μl;40IU/ml)的Neurobasal/2%B27替換,並在37℃的CO2培養箱中培養5分鐘。培養後,將含有肽或穀氨酸受體阻礙劑(MK801/CNQX)的Neurobasal/2%B27添加到培養孔中,在37℃的CO2培養箱中進一步培養10min。培養後,去除孔中的培養基並用100μl含穀氨酸(100μM)的Neurobasal/2%B27培養基替換,隨後的處理如上所述。對於所有的實驗,採用穀氨酸處理的無肝素處理肽對照經過相同的培養步驟並採用相同的培養基添加物。體外局部缺血/OGD模型和肽培養用於原代皮質神經元培養的體外局部缺血模型的建立如現有技術所描述(Meloni等,2011)。簡單的說,去除孔中(玻璃96-孔板式)的培養基,並用315μl無葡萄糖平衡鹽溶液(BSS;116mMNaCl、5.4mMKCl、1.8mMCaCl2、0.8mMMgSO4、1mMNaH2PO4;pH6.9)衝洗,然後添加60μl的含穿膜肽或對照肽(見表1)的BSS。還包括由穀氨酸受體阻滯劑(5μMMK801/5μM6-氰基-7-硝基喹喔啉:MK801/CNQX)組成的非肽陽性對照。體外缺血是通過將孔板置於37℃的厭氧培養箱(DonWhitelyScientific,英國;5%CO2、10%H2和85%氬氣的氣氛,98%的溼度)中培養55分鐘而引發的。從厭氧培養箱取出後,將60μl的Neurobasal/2%N2補充物添加到孔中,培養物進一步在37℃、CO2培養箱中孵化培養24h。在37℃的CO2培養箱中培養前,對照培養物採用同樣的BSS清洗步驟並採用相同的培養基添加物作為缺血性治療培養物。對於OGD暴露前實驗,其步驟與在穀氨酸模型中所描述的一樣,除了10分鐘肽預處理是利用100μlNeurobasal/2%B27培養基進行的。對照培養物經歷同樣的BSS清洗步驟並採用相同的培養基添加物作為OGD治療培養物。體外缺血模型也可用於bEND3細胞、SH-SY5Y細胞和星形膠質細胞。對於bEND3細胞,厭氧培養延長至2-3小時,從厭氧培養箱中移出後,緊接著將60μlDMEM/2%FCS加入到孔中。對於SH-SY5Y細胞,省去第一次BSS清洗步驟(315μl),厭氧培養延長到2-5小時。從厭氧培養箱中移出後,緊接著將60μl的DMEM/2%FCS培養液加入到孔中。對於星形膠質細胞,厭氧培養延長到1:15至2:00小時,從厭氧培養箱移出後,緊接著將60μl的DMEM/2%FCS培養液加入到孔中。體外神經元、bEND3和星形膠質細胞毒性模型和肽培養。對於神經元培養物,通過移除培養基和添加100μl的50%Neurobasal/2%N2補充物和含CPPs、JNKI-1D-TAT或TAT-NR2B9c的50%BSS,將肽加入到培養孔(96孔板式)。對照培養物僅加入50%Neurobasal/2%N2補充物和50%BSS培養基。培養物在37℃的CO2培養箱中培養20小時,然後使用MTS法評估細胞存活率。對於bEND3培養物,通過移除培養基和添加100μl含肽的DMEM/2%FCS,將肽加入到培養孔中。對照培養物只添加100μl的DMEM/2%FCS。在37℃的CO2培養箱中培養0.5、1或2小時,然後使用MTS法評估細胞存活率。對於星形膠質細胞培養物,通過移除培養基和加入100μl含肽的DMEM/2%FCS,將肽加入到培養孔(96孔板式)中。對照培養物只添加100μl的DMEM/2%FCS。在37℃的CO2培養箱中培養24小時,然後使用MTS法評估細胞存活率。細胞存活率評估和統計分析在刺激後的24小時,通過光顯微鏡檢測神經元培養物,定性評估神經細胞存活率。通過3-(4,5,二甲基噻唑-2-基)-5-(3-羧甲氧基-苯基)-2-(4-磺苯基)-2H-四唑鹽(MTS)法(Promega,澳大利亞)定量檢測神經細胞存活率。MTS法測量四唑鹽的細胞轉化為水溶性的棕色甲瓚鹽,並在490nm下進行分光光度法檢測。MTS吸光度值可轉換成與未處理和已處理對照有關的細胞存活率比例,未處理對照作為100%存活率,並作為平均±SEM。對於使用星形膠質細胞、bEND3和SH-Y5Y細胞的研究,原始的MTS值用於作圖。通過ANOVA分析存活率數據,然後進行post-hocFischer’sPLSD檢驗,P<0.05認為有統計學顯著性。所有的試驗中使用到4-6孔。大鼠永久局灶性腦缺血模型-實驗組和治療所有治療採用隨機的並採用盲法方式給藥。在MCAO後30分鐘施用肽(DR9:rrrrrrrrr-NH2;R12、R15、R18、硫酸魚精蛋白;魚精蛋白)或媒介物(鹽水:0.9%氯化鈉)處理溶液。包含DR9鹽水的肽治療經由右頸內靜脈注射5-6分鐘提供1μmol/kg或1000nmol/kg的靜脈負荷劑量。大鼠永久性局灶性腦缺血模型這項研究是經西澳大利亞大學動物倫理委員會(AnimalEthicsCommitteeoftheUniversityofWesternAustralia)批准的。體重為270克到350克的雄性SpragueDawley大鼠,在12小時光照-黑暗周期循環的可控居住環境下,自由進食和飲水。實驗動物禁食過夜,並遭受永久大腦動脈阻塞(MCAO),如下所示。通過4%異氟醚以及N2O和O2按2:1混合掩面引發麻醉。麻醉維持在1.7-2%異氟醚。採用雷射都卜勒血流儀連續監控(血流量計,AD儀器,澳大利亞雪梨)腦血流量(CBF)。探針位於前囪的1毫米尾側和4毫米外側(右)。插管插入右股動脈中以連續監測血壓並提供血糖和血液氣體讀數樣本。採用血糖儀(MediSenseProducts,AbbottLaboratories,Bedford,MA,美國)測定血糖,並採用血氣分析儀(ABL5,Radiometer,丹麥哥本哈根)測定血氣。血壓保持在80-100毫米汞柱。在手術過程中,直腸溫度保持在37±0.5℃,必要時用暖風機加熱。對於靜脈輸注,將含肝素的生理鹽水泵入PVC管中,並固定在右頸內靜脈合適的位置,然後通過背中期肩胛骨切口外化到繫繩/旋轉系統(InstechLaboratories,美國費城),設計該系統允許自由移動。右側頸總動脈(CCA)經腹頸部切口暴露。經甲狀腺上動脈和枕動脈電療後,分離出頸外動脈(ECA)。結紮和電療ECA的分離部分以形成殘枝。移除頸動脈體並結紮翼顎動脈。具有0.39毫米直徑矽接點的4-0尼龍單絲(Doccol,Redlands,CA,USA)通過ECA殘枝插入到CCA中,平臥進入頸內動脈(ICA)直到雷射都卜勒血流儀記錄腦血流量的基線下降>30%。在剩下的實驗中,單絲固定在兩個地方(ECA殘枝的底部和ICA上)。在動物的頭部和腿部切口處給予由哌替啶(肌肉注射3mg/kg)和布比卡因術後鎮痛(皮下注射1.5mg/kg)組成的術後止痛劑。手術後動物置入氣候控制室進行恢復,必要時,通過冷/熱風扇3-4小時,監測並維持它們的核心體溫在37℃±0.5℃。組織處理與梗死體積測量在MCAO後24h通過腹腔內注射戊巴比妥鈉(900mg/kg)處死動物。安樂死後,移除大腦並放置在0.9%氯化鈉的無菌容器中,然後放置在-80℃冰箱中7分鐘。然後從大腦與小腦交界處到12毫米喙點冠狀切除2毫米厚的切片。切片立即用1%的2,3,5-氯化三苯基四氮唑(TTC,Sigma,聖路易,MO,USA)在37℃下染色20分鐘,然後在室溫下用4%福馬林固色至少18-24小時,測量腦梗死體積。掃描切片以及操作者無視治療狀況使用ImageJ第3版(NIH,USA)分析圖像。總的腦梗死體積是由在2毫米切片的兩邊增加梗塞組織區域決定的。這些測量的區域乘以切片厚度的一半(1毫米),通過乘以影響正常半球面積的比來影響正腦水腫。統計分析對於腦梗死體積測量,通過t-檢驗(R9D試驗)或方差分析(ANOVA)比較肽治療組和載體對照組,然後進行post-hocFischer’sPLSD檢驗(R12、R15、R18和Ptm試驗)。實驗例1暴露於穀氨酸後神經保護功效CPPsTAT-D、R9和穿膜肽以劑量響應方式提供重要神經保護功效(圖1a,表3)。目測評估損傷後培養物也證實神經保護功效,其神經元存活率範圍從未經穀氨酸暴露處理的培養物的約5%到R9處理的培養物的100%存活。R9是最有效的肽,具有0.78μM的IC50值,其次是穿膜肽(IC50:3.4μM)和TAT-D(IC50:13.9μM)。Pep-1肽是無效的。穀氨酸受體阻滯劑和對照肽(JNKI-1D-TAT、PYC36L-TAT)在這個模型中(圖1a)中也是高效的。此外,TAT-D肽顯示與TAT-L-肽相似水平的神經保護功效(圖1b)。暴露於穀氨酸前,將CPPs洗淨,只有R9(在這個實驗中)顯示出高水平的神經保護功效(圖1c)。在暴露於穀氨酸後立即加入R9和R12肽,R9和R12肽也是高效的,在刺激後15分鐘加入R9,是中度有效的。相反,暴露於穀氨酸後立即加入或15分鐘後加入JNKI-1D-TAT和NR29c(也稱為TAT-NR2B9c)肽,沒有顯著增加神經元的存活。圖1e-j顯示肽R1、R3、R6、R9、R12、R15、R18、R9/E9(也稱為E9/R9)、R9/tPA/R9(也稱為R9/X7/R9)以及對照肽JNKI-1D-TAT和NR29c在穀氨酸模型中具有額外功效的數據。在穀氨酸模型中使用R1、R3、R6、R7、R8、R9、R12、R15和R18的劑量響應研究中顯示:i)R1、R3、R6和R7顯示出沒有至微小的神經保護功效;ii)5μM的R8顯示出神經保護功效,iii)其他肽的效價順序是R15>R18>R12>R9;iv)R15和R18的神經保護功效在高濃度(5μM)降低;見表4,圖1e、1l、1j、6、7和8。在劑量響應研究中,在穀氨酸模型中使用R9、DAHK(白蛋白的最後四個以N-端胺基酸結尾)PTD4(具有增加33倍的細胞穿透能力的改性TAT肽)(Ho等,2001年),R9/E9(Arg-9/Glu-9;中性肽)和R9/tPA/R9(R9位於組織纖溶酶原激活劑酶肽切割位點的側面)表明:i)PTD4和R9/E9顯示出沒有至微小的神經保護功效;DAHK具有低水平的神經保護功效;和ii)R9/tPA/R9具有R12和R18之間的效力。並且TAT-NR29C肽(C-端NR2BNMDRA受體亞基肽;阻斷NMDRA受體和PSD95蛋白質的通信以阻斷NO(一氧化二氮)生產;Aarts等,2002)是無效的。見表4和圖1d、g、h。在穀氨酸模型中,肽R12比R9更有效(根據0.625μM濃度的神經保護功效),與PCY36-TAT相比,肽R12和R9都更有效,而NR29c是無效的(圖1g)。肽R12比R9更有效(根據濃度在0.5μM和1μM的神經保護功效),而R1、R3、R6和NR29c是無效的(圖1i)。兩個不同的公司合成的肽R9在穀氨酸模型中也有相類似的效力(圖1j)。肽R9、R12、R15和R18在穀氨酸刺激僅5min時加入到神經元培養物中(圖28);或肽R9、R12、R15和R18在神經元培養物預暴露於穀氨酸10min後在刺激前從培養中移出(圖29),肽R9、R12、R15和R18是有效的。在穀氨酸刺激前,肽R12和R15加入到神經元培養10分鐘、1到4小時,肽R12和R15是有效(圖30)。在穀氨酸模型中,肽PTD4顯示低水平的神經保護功效,肽E9/R9顯示無保護作用(圖33),而富含精氨酸的肽XIP和NCXBP3表現出高水平的保護作用(圖34、35)。聚賴氨酸-10肽(K10)和TAT-NR2B9c肽分別在穀氨酸模型(圖36)和NMDA模型(圖37)中顯示低水平的神經保護功效。在穀氨酸模型中,R8和R9融合到鈣蛋白酶切割位點(Cal/R9)顯示中等到高水平的神經保護功效(圖38)。圖39和40顯示在穀氨酸模型中負電荷分子肝素阻斷R9D、R12、R15和PYC36-TAT的神經保護功效,但不影響穀氨酸受體拮抗劑(5μMK801/5μMCNQX)。圖41顯示在穀氨酸模型中,當JNKI-1肽融合到(非精氨酸)kFGFCPP,JNKI-1肽不依賴於內吞作用攝取,不具有神經保護功效,kFGF肽也是無效的。相反,TAT-JNKI-1和JNKI-1-TATD具有神經保護功效。圖44顯示穀氨酸處理後,在神經元培養物中,肽R9D、R12、R15和PYC36-TAT、TAT、TAT-NR2B9c、TAT-JNKI-1和JNKI-1-TATD和穀氨酸受體阻滯劑(5μMK801/5μMCNQX)不同程度的減少鈣內流。實驗例2紅藻氨酸暴露後的神經保護功效暴露於紅藻氨酸後,TAT-D、R9和穿膜肽具有神經保護功效,但比在穀氨酸模型中的效力差,並且不總是顯示出典型的劑量響應模式(圖2,表2)。Pep-1是無效的。R9是最有效力的肽,神經元存活從約20%增加到最大值約80%。R9、穿膜肽和TAT-D的IC50值分別為0.81、2.0及6.2μM。在這個模型中,穀氨酸受體阻滯劑、JNKI-1D-TAT和PYC36L-TAT也是有效的(圖2)。實驗例3體外局部缺血/氧糖剝奪(OGD)後的神經保護功效以下體外缺血後,四個CPPs都顯示出神經保護功效(圖3,表2)。Arg-9(IC50:6.0μM)和TAT-D(IC50:7.1μM)的神經保護功效相似;劑量響應模式後的效力和神經元存活率從約10%增至40-50%。隨著穿膜肽濃度(≥5μM)的增加,神經保護功效消失,而Pep-1是唯一在較低濃度(1-5μM)具有神經保護功效。在此模型中,穀氨酸受體阻滯劑和PYC36L-TAT也是有效的(圖3a)。此外,當體外局部缺血後,加入R9、R12、R15和R18顯示神經保護功效,但較高濃度的R15和R18功效降低(圖3b)。當暴露於體外局部缺血中時,R9也減少了bEND3和SH-5YSY細胞死亡(圖3c)。當暴露體局部外缺血時,R18也減少了星形膠質細胞死亡(圖42)。圖6顯示在穀氨酸模型中,使用R9、R10、R11、R12、R13和R14的劑量響應研究,表明所有肽的濃度在1和5μM之間具有明顯的神經保護功效,除了R11,其在濃度2和5μM具有明顯的神經保護功效。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。圖7顯示在穀氨酸模型中,使用R9D、R13、R14和R15的劑量響應研究,表明了所有肽在濃度為1至5μM之間具有神經保護功效。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。如圖8所示,在穀氨酸模型中,使用R6、R7和R8、R9的劑量響應研究表明,肽R8和R9分別在濃度5μM和1μM和5μM顯示出明顯的神經保護功效,而R6和R7沒有顯示明顯的神經保護功效。均值±SEM:N=4;*P<0.05。(肽濃度以μM表示)。圖31顯示使用R9、R12、R15和R18的劑量響應研究,在OGD後加入肽培養15min。R12、R15和R18在濃度為1μM和5μM時具有神經保護功效,但R9在1μM和5μM沒有神經保護功效。在OGD之前,肽R12和R18加入到神經元培養物中10分鐘、1至3小時,肽R12和R18是有效的(圖32)。表3:三個損傷模型中細胞穿膜肽和對照肽的IC50值*圖1a、圖2和圖3a示出了基於劑量的響應圖。#JNKI-1D-TAT和PYC36L-TAT肽的IC50值來自Meade等,(2010a,b)。N/A=不適用,因為肽在較高劑量是無效的或增加了細胞死亡。-=未獲得數據。表4:穀氨酸模型*中聚精氨酸肽的IC50值*圖1e示出了基於劑量響應圖。N/A=不適用,因為肽在最高濃度(5μM)實驗中無效或效果很小。實驗例4動物試驗在已經進行的初期動物試驗中,表明Arg-9(R9)、R18和魚精蛋白(Ptm)在體內具有神經保護活性。這些試驗表明在大鼠永久大腦中動脈阻塞(MCAO)中風模型中R9D肽的功效。R9D肽是在MCAO後30分鐘進行靜脈注射施用。測量MCAO後24h腦梗死體積(腦損傷)(均值±SEM)。從圖5和圖27可以看出,(Ns=每組8-12隻動物)經R9D、R18、魚精蛋白(Ptm)治療減少MCAO中風後腦梗死體積(腦損傷)約20%,表明具有統計學顯著性的神經保護功效。一般性意見和討論申請人對皮層神經元培養物暴露於穀氨酸、紅藻氨酸或體外局部缺血(氧糖剝奪)後,作為已知的神經保護功效例子的TAT以及其他3種CPPs(穿膜肽、R9和Pep-1)的神經保護性能進行評價。此外,在使用體外缺血模型在星形膠質細胞、腦內皮細胞系(bEND3)和/或神經母細胞瘤細胞系(SH-SY5Y)中對聚精氨酸肽(R9、R12、R15、R18)和/或富含精氨酸的魚精蛋白肽進行評價。R9、穿膜肽和TAT-D在穀氨酸(IC50:0.78、3.4、13.9μM)損傷模型和紅藻氨酸(IC50:0.81、2.0、6.2μM)損傷模型中顯示出一致的和高水平的神經保護活性,而Pep-1是無效的。在穀氨酸模型中TAT-D同種型與TAT-L同種型具有相似的功效。在神經元暴露於穀氨酸前洗淨,R9表現出功效。然而,R9比起之前已經被證明具有神經保護功效的肽,具有更為明顯的功效,即TAT-D、TAT-L、PYC36L-TAT和JNKI-1D-TAT。在體外局部缺血後神經保護功效是更多變的。所有肽提供某種程度的神經保護功效(IC50;R9:6.0μM,TAT-D:7.1μM,穿膜肽/Pep-1:>10μM)。陽性對照肽JNKI-1D-TAT(JNK抑制肽)和/或PYC36L-TAT(AP-1抑制肽)在所有模型中具有神經保護功效。在穀氨酸治療實驗後,在刺激後立即加入R9是高效的,在刺激後15分鐘加入R9是中度有效的,而在刺激後加入JNKI-1D-TAT對照肽是無效的。在已進行的初期動物試驗中,表明R9、R12、R18和魚精蛋白在體內具有神經保護活性。在穀氨酸模型中,使用R1、R3、R6、R9、R12、R15和R18的劑量反應研究表明:i)R1、R3、R6、R7顯示出神經保護功效;ii)其他肽的效價順序是R15>R18>R12>R9;和iii)R15和R18在高濃度(5μM)測試時神經保護功效降低。在穀氨酸模型中,使用R9、DAHK(白蛋白的最後四個以N-端胺基酸結尾)PTD4(具有增加33倍的細胞穿透能力的改性TAT肽;Ho等,2001年),R9/E9(Arg-9/Glu-9;中性肽)和R9/tPA/R9(R9位於組織纖溶酶原激活劑酶肽切割位點的側面)劑量響應研究表明:i)PTD4和R9/E9顯示出神經保護功效;DAHK具有低水平的神經保護功效;和ii)R9/tPA/R9具有R12和R18之間的效力。並且TAT-NR2B9C肽(C-端NR2BNMDRA受體亞基肽阻斷NMDRA受體和PSD95蛋白質的通信,以阻斷NO(一氧化二氮)生產;Aarts等2002)。聚精氨酸和富精氨酸的(魚精蛋白)肽也能降低體外局部缺血後星形膠質細胞、bEND3和SH-SY5Y細胞的死亡。這些研究結果表明,本發明的肽具有抑制與興奮性和缺血性損傷有關的神經破壞事件/途徑的能力。具有≥9個精氨酸胺基酸殘基的聚精氨酸肽尤其具有神經保護功效。本發明的肽的細胞保護性質表明它們是損傷後遞送神經保護藥物到CNS的理想分子載體和/或它們本身作為潛在的神經保護劑。因此本發明的肽在不同體外損傷模型中表現出神經保護性質,已經表明所述體外損傷模型能夠轉換成體內模型。在進行的初始動物試驗中神經保護功效進一步加強。R9優越的神經保護功能是意想不到的;在穀氨酸模型和紅藻氨酸模型中R9的IC50值分別比TAT-9強17倍和7倍,是唯一一種甚至在神經元暴露於穀氨酸前洗淨仍有神經保護功效的肽。這一發現表明增加精氨酸殘基和/或略高的R9淨電荷(10vs9,pH=7)是興奮性毒性後的神經保護功效的重要因素。此外,在神經元培暴露於穀氨酸前衝洗,除了R9,TAT和穿膜肽的效力損失的確切原因還不清楚,這可能和R9胞內吸收速度有關,而不是胞外機制。這得到以下發現的支持,暴露於穀氨酸後加入R9是有效的,而JNKI-1D-TAT肽是無效的。此外,當在穀氨酸損傷模型中對精氨酸和聚精氨酸肽R1、R3、R6、R9、R12、R15和R18進行評估,只有肽R9、R12、R15和R18在劑量測試中具有顯著神經保護功效;R15看起來是最有效的肽。含tPA裂解連接位點的雜交R9肽(R9/tPA/R9)在穀氨酸模型中也是高效的。當在暴露於穀氨酸後加入,R9也比R12更有效。有趣的是,PTA4(具有增加33倍的細胞穿透能力的改性TAT肽,;Ho等,2001年),R9/E9混合物和NR29c(阻斷NMDA/穀氨酸受體誘發NO生成的TAT融合肽)和DAHK(白蛋白最後四個以N-端胺基酸結尾),在暴露於穀氨酸後很大程度上無效。在體外缺血模型中,厭氧培養(即損傷再灌注階段中)後加入肽R9、R12、R15和R18也是有效的。體外局部缺血後,R9也能夠保護腦內皮細胞(bEND3細胞)和神經母細胞瘤細胞(SH-5YSY)。如上文所述,研究的結果證明,穿膜肽和Pep-1也具有神經保護性質。穿膜肽和Pep-1肽相互之間或與TAT/R9肽沒有胺基酸序列相關性。有趣的是穿膜肽在興奮毒性模型中具有高神經保護功效(IC50s:3.4和2μM),但在體外缺血模型中效力降低,隨著濃度的增加效力降低。Pep-1肽在興奮性模型中通常無效並且在一些實驗中,似乎會增加神經元死亡(數據未顯示),但在體外缺血後低濃度下表現出神經保護功效。有趣的是,在神經元培養物暴露於穀氨酸之前從神經元培養物中洗淨穿膜肽,目測觀察顯示一些早期的神經保護功效(數據未顯示)。因此,在損傷模型中,穿膜肽和Pep-1行為互相不同,並且和TAT/R9肽不同。在興奮性和缺血性損傷模型中,四種CPPs不同的神經保護響應可能與多肽的理化性質有關,更具體地說是與它們的誘導內吞性質有關。此外,有可能的是,CPPs的神經保護功效是在細胞膜(如受體、離子通道)進行介導的。Xu等(2008)認為TAT可能改變細胞膜,從而影響細胞表面受體的功能(比如NMDA受體),使得鈣內流減少。然而,可以預見的是,本發明的肽或多種肽能夠阻止NMDA受體作用和/或阻止、抑制或減慢鈣的內流。替代機制是CPPs與外線粒體膜相互作用並穩定外線粒體膜,從而有助於保持線粒體功能。潛在的益處是維持ATP合成、減少活性氧產生和提高鈣處理。為此,申請人已經觀察到Arg-9可以提高正常神經元和損傷後MTS吸光度水平並超過基線水平(如圖1A、15μM)。由於MTS對甲瓚產物的減少主要發生在線粒體中,Arg-9增強甲瓚水平的能力,有利於肽改善線粒體功能。尤其涉及R9和TAT的另一種潛在機制是這些富精氨酸肽抑制鈣依賴性前蛋白轉化酶弗林蛋白酶(Kacprzak等,2004),從而阻止潛在破壞性的蛋白質的激活。本發明證明富含陽離子胺基酸的CPP,尤其是富含精氨酸的CPPs或載體-肽(例如R12、R15、魚精蛋白),相對於CPPs,大體上顯示出高水平的神經保護功效。這就提出了一種可能性,融合到CPP的神經保護肽的作用機制很有可能的是,如果不是完全是,載體-肽的增強的神經保護功效的結果。此外,其中富含精氨酸的CPPs發揮它們神經保護功效的機制可能與細胞內吞作用(主要的載體肽細胞攝取路線)有關,而不是通過與具體的細胞質靶點相互作用。與此相反的是,融和到載體-肽的神保護肽,通過內吞作用進入細胞,在神經肽可以與其細胞質靶點相互作用之前,必須首先逃脫核內體,這是一個眾所周知的非常無效的過程(Al-Taei等,2006;El-Sayed等2009;Appelbaum等,2012;Qian等,2014),因此致使肽很可能不能通過與其預定目標作用而起作用。關於CPP細胞內進入,主要機制被認為是通過內吞作用(巨胞飲)(Palm-Apergi等,2012)。雖然與本發明不太相關,最近一份報告已證明運載貨物(cargo)的性質也可通過某種CPPs促進直接進入細胞機制(Hirose等,2012)。然而,可能高度相關的是具體物體、肽或其他物質是如何通過加強其神經保護功效、提高轉運效率和/或Cardozo等(2007)所證明的增加它們的毒性來影響CPPs。特別重要的是,因為如上文所述,運載貨物本身具有神經保護,這使得識別CPP和運載貨物之間的神經保護功效是非常困難的。例如,現有研究(Meade等,2010a),在穀氨酸模型中從PYC36肽到TAT-D肽(AM8D-TAT)添加三個胺基酸殘基(Pro、Lys、Ile)導致IC50值從TAT-D的>15μM降到AM8D-TAT的1.1μM。TAT肽對照的正面效果並不總是可以觀察到的。存在很多可能的解釋以及處理這個問題:首先必須區分僅使用TAT肽(即GRKKRRQRRRG)的研究與使用融和到報導蛋白質或肽(例如透明質酸(HA)和/或6倍組氨酸標記,雜交肽)的TAT作為對照的研究(如綠色螢光蛋白(GFP)、β-半乳糖)。關於使用TAT肽本身作為對照的研究,在使用的劑量下和/或損傷模型太嚴重不能顯示TAT的神經保護功效,TAT有可能是無效的。例如,Boresello等,(2003年)在皮層神經元培養物暴露於100μMNMDA12、24或48小時後,並未檢測到TAT肽具有神經保護功效。與此相反,L-JNKI-1肽在12和24小時有效,而蛋白酶抵抗D-JNKI-1肽在所有的時間點均有效。考慮到JNKI-1肽與TAT肽相比,JNKI-1肽具有優越的功效,濃度測試時,TAT有可能無神經保護功效或由於NMDA刺激嚴重性任何神經保護功效都被壓制。Ashpole和Hudmon(2011年)研究發現,皮質神經元培養物暴露於穀氨酸後,觀察到TAT肽的適度保護作用。此外,作者推斷由於TAT肽幾乎沒有保護作用,觀察CAMKII抑制肽的神經保護功效並不是因為"導入序列"(即TAT)。不過,也不排除CAMKII抑制肽增加TAT肽的效價。最後,TAT肽可能在具體的損傷模型和細胞類型中才具有神經保護性質。在使用融和到報導蛋白或對照肽的TAT研究中,除了以上提出的幾點,對照蛋白/肽也有可能抑制或抵消TAT肽的神經保護性能。基於許多已經使用融合有蛋白質/多肽的TAT作為對照並顯示不具有神經保護功效的研究,情況似乎是這樣的(例如Kilic等,2003;Doeppner等,2009)。然而,需要記住,蛋白質是CPP是不爭事實,但並不意味著它將具有神經保護功效,這可通過PTD-4肽(具有比TAT本身好33倍的轉導效率的改性TAT肽;Ho等,2001年)幾乎不具有神經保護性能這一事實進行證明。表5:不同的魚精蛋白(鮭魚)肽的胺基酸序列;胺基酸殘基/精氨酸殘基和分子量。(魚精蛋白肽序列(鮭魚)存在於硫酸魚精蛋白中,用於臨床使用或者SwissProt資料庫。[a=硫酸魚精蛋白1的HPLC檢測到的峰1–4;b=SwissProt資料庫中的序列(Swissprot:P14402)。(1)HoffmannJA1,ChanceRE,JohnsonMG.1990.Purificationandanalysisofthemajorcomponentsofchumsalmonprotaminecontainedininsulinformulationsusinghigh-performanceliquidchromatography.(採用高效液相色譜法純化與分析含有大麻哈魚魚精蛋白的胰島素劑的主要成分),ProteinExprPurif.1(2):127-33.(2)ChangLC,LeeHF,YangZ,YangVC.2001.Lowmolecularweightprotamine(LMWP)asnontoxicheparin/lowmolecularweightheparinantidote(I):preparationandcharacterization(作為無毒肝素的低分子量魚精蛋白(LMWP)/低分子量肝素解毒劑).PharmSci.3(3):E17).實驗例1暴露於穀氨酸後的神經保護功效具有硫酸魚精蛋白(魚精蛋白;Ptm)具有重要神經保護功效存在劑量響應關係(圖9、10、12、13、15)。損傷後培養的目測評估也證實硫酸魚精蛋白的神經保護功效,從未處理的暴露於穀氨酸的培養物約5%存活率到魚精蛋白處理的培養物85-100%存活率之間。此外,預暴露於魚精蛋白5或10分鐘也具有高神經保護功效,使得100%神經元存活(圖11)。在劑量響應試驗中,低分子量魚精蛋白(LMWP)、魚精蛋白1(Ptm1)、魚精蛋白2(Ptm2)、魚精蛋白3(Ptm3)、魚精蛋白4(Ptm4)、魚精蛋白5(Ptm5)、肽均具有神經保護功效(圖12、13、15)。在預暴露於魚精蛋白和LMWP的試驗中,當神經元在穀氨酸刺激前立即以及穀氨酸刺激前1或2小時暴露於魚精蛋白中,魚精蛋白具有神經保護功效,而僅在穀氨酸刺激前立即暴露於LMWP中,LMWP具有神經保護功效(圖14)。實驗例2氧糖剝奪(OGD)後的神經保護功效在OGD模型中,在OGD刺激前1小時(圖18)或OGD刺激後(圖16、17),用魚精蛋白肽處理神經元,魚精蛋白具有神經保護功效。此外,OGD後加入魚精蛋白15min或30min也具有神經保護功效(圖19、20、21、22)。神經元在OGD前1小時預暴露於LMWP,LMWP不具有神經保護功效,但OGD後加入LWMP15分鐘,具有神經保護功效。此外在OGD後加入魚精蛋白肽(Ptm1-Ptm5)和LMWP15分鐘,肽Ptm2、Ptm4、Ptm5和LMWP均具有神經保護功效(圖22)。實驗例3氧糖剝奪(OGD)後對bEND3細胞的保護在OGD模型中,在OGD前15min用魚精蛋白、魚精蛋白4(ptm4)處理血腦屏障(bEND3)內皮細胞時,上述肽對血腦屏障(bEND3)內皮細胞有保護作用(圖23、24)。此外,根據MTS法檢驗,bEND3暴露於濃度範圍從1.25-15μM的魚精蛋白中0.5-2小時,不會產生顯著性的毒性(圖25、26)。圖43顯示暴露24小時後,濃度從2.5到10μM的Ptm1-Ptm5和低分子量魚精蛋白(LMWP)沒有引起星形膠質細胞顯著地死亡。一般意見和討論申請人評估了在皮質神經元培養物暴露於穀氨酸後或體外局部腦缺血後,富含精氨酸的硫酸魚精蛋白(魚精蛋白;Ptm),魚精蛋白肽1-5(Ptm1-Ptm5,SEQ.ID.NOs分別為32到36),和低分子量魚精蛋白(LMWP,SEQ.ID.NO.37)對皮質神經元的神經保護性能。這兩種損傷模型通常用於模仿缺血性腦中風的效果。申請人還評估了魚精蛋白肽保護血腦屏障(bEND3)內皮細胞免於OGD的應用。魚精蛋白在穀氨酸和OGD損傷模型中都顯示出一致的和高水平的神經保護功效,同時魚精蛋白bEND3細胞中也提供保護。LMWP在神經元穀氨酸和OGD損傷模型中的效果略差(根據劑量濃度以達到與魚精蛋白等效的神經保護功效)。這最可能是由於LMWP肽幾乎不含精氨酸殘基。魚精蛋白肽1-5(Ptm1-Ptm5)在穀氨酸模型中也具有高效的神經保護功效,而Ptm2、Ptm4和Ptm5在OGD模型中具有神經保護功效。在使用bEND3細胞的OGD研究中,在不同的OGD期間後(2h:15min、2h:30min或2h:45min),bEND3細胞預暴露於魚精蛋白15分鐘,顯著增加細胞存活率,從而起保護作用。此外,在OGD模型中,預暴露於Ptm415分鐘,也可顯著增加細胞存活率,從而起保護作用。在bEND3毒性研究中,使用不同濃度的魚精蛋白,表明魚精蛋白即使濃度高達15μM,其不具有毒性。這些研究結果表明,本發明的魚精蛋白肽具有抑制與興奮性和缺血性損傷有關的神經破壞事件/途徑的能力。並且,由於在預曝光試驗中魚精蛋白的效果,一個新的重要發現是在神經元暴露於穀氨酸或OGD前1到2小時,魚精蛋白處理神經元可以通過減少細胞死亡引起神經保護響應。這是重要的,因為有大量的腦血管(例如頸動脈內膜切除手術)和心血管(如冠狀動脈旁路移植術)手術治療中,患者有可能遭受缺血性腦損傷或中風的風險,造成的腦損傷。因此,在手術前的1-2小時施用魚精蛋白肽以保護大腦不遭受任何此類腦缺血性事件。本發明的肽的細胞保護性能顯示它們是治療CNS損傷的理想的神經保護藥物。此外,它們還可能具有細胞穿透性能[魚精蛋白是FDA批准用於基因治療載體(DNA,病毒載體;Sorgi等,1997)和LMWP作為細胞穿膜肽使用;Park等,2005],它們是將神經保護藥物運輸到到損傷後的CNS的理想載體分子。本發明的肽的神經保護功效很可能與肽的理化性質有關。此外,富含精氨酸的魚精蛋白肽的神經保護活動可能是在細胞膜(如受體、離子通道)介導的。研究表明,富含精氨酸的肽包括TAT(Xu等,2008)、R6(Ferrer-Montiel等,1988)和R9-CBD3(Feldman和Khanna2013)和TAT-CBD3(Brustovetsky等,2014)可能會影響細胞表面受體和離子通道的功能(如NMDA受體),使得鈣內流減少。然而,可以預期的是,本發明中肽或多種肽可以阻斷NMDA受體功能和/或阻斷、下調或減少鈣內流。一種替代機制是魚精蛋白肽與線粒體外膜相互作用和穩定線粒體外膜,從而有助保持線粒體功能。潛在的益處是維持ATP合成、減少活性氧產生和提高鈣處理。為此目的,申請人已觀察到,在正常神經細胞中以及損傷後,魚精蛋白可以提高MTS吸光度水平高於基線水平(如圖11,12;5μM)。由於MTS減少甲瓚產物主要發生在線粒體中,魚精蛋白增加甲瓚水平的能力可以作為肽改善線粒體功能的支持。另一個潛在的機制是這些富含精氨酸多肽抑制鈣依賴性前蛋白轉化酶酶弗林蛋白酶(Kacprzak等,2004),並因此阻止激活潛在破壞蛋白質。對於魚精蛋白肽的細胞內進入,富含精氨酸的肽的主要機制被認為是通過內吞作用(巨胞飲)(Palm-Apergi等,2012)。因此可能的是,跨質膜的過程中肽的內吞作用導致細胞表面結構的內含體內在化(見圖46)。在建立神經元興奮性毒性和局部缺血模型中,下調離子通道可能有利於降低正常的鈣和其它離子的神經損害性內流。申請人認為他們已經發現一類或一組新的肽,可以作為CPPs,以及作為神經保護肽。申請人已發現聚精氨酸或富含精氨酸的肽,尤其是那些選自魚精蛋白或低分子量魚精蛋白(和混合物及其衍生物,尤其是以市售的的魚精蛋白硫酸鹽的形式)具有獨特的神經保護或神經活性的性能。有證據表明,本文中公開的肽作為本發明的一部分,在應用於體內(Vaslin等,2009)和用於治療神經損傷,具有相同範圍的神經保護或神經活性的性能。申請人因此發現,富含精氨酸的肽和與TAT無關的CPPs具有獨特的神經保護性能,尤其是聚精氨酸序列和長度為10和32個胺基酸的序列具有超過10個精氨酸殘基(如魚精蛋白、LMWP及其衍生物)的序列。證據顯示,本發明的CPPs在應用於體內和用於治療神經損傷時,具有相同範圍神經保護性能。參考文獻AartsM,LiuY,LiuL,BesshohS,ArundineM,GurdJW,WangYT,SalterMW,TymianskiM(2002)TreatmentofischemicbraindamagebyperturbingNMDAreceptor-PSD-95proteininteractions.Science298:846-850.ARAMIS(2011)Projectnumber:8909.1.http://www.aramis.admin.ch.ArthurPG,MatichGP,PangWW,YuDY,BogoyevitchMA(2007)Necroticdeathofneuronsfollowinganexcitotoxicinsultispreventedbyapeptideinhibitorofc-junN-terminalkinase.JNeurochem102:65-76.BorselloT,ClarkePG,HirtL,VercelliA,RepiciM,SchorderetDF,BogousslavskyJ,BonnyC(2003)Apeptideinhibitorofc-JunN-terminalkinaseprotectsagainstexcitotoxicityandcerebralischemia.NatMed9:1180-1186.BrugidouJ,LegrandC,MéryJ,RabiéA(1995)Theretro-inversoformofahomeobox-derivedshortpeptideisrapidlyinternalisedbyculturednaurones:anewbasisforanefficientintracellulardeliverysystem.BiochemBiophysResCommun214:685-693.ColomboA,ReplciM,PesaresiM,SantambrogioS,ForloniG,BorselloT(2007)TheTAT-JNKinhibitorpeptideinterfereswithbetaamyloidproteinstability.CellDeathDiffer14:1845-1848.CraigAJ,MeloniBP,WattPM,KnuckeyNW(2011)AttenuationofneuronaldeathbypeptideinhibitorsofAP-1activationinacuteanddelayedinvitroischaemia(oxygen/glucosedeprivation)models.IntJPeptResTher17:1-6.DoeppnerTR,NagelF,DietzGPH,WeiseJ,L,SchwartingS,M(2009)TAT-HSP70-mediatedneuroprotectionandincreasedsurvivalofneuronalprecursorcellsafterfocalcerebralischemiainmice.JCerebBloodFlowMetab29:1187-1196.DolginE(2012)Toserveandneuroprotect.NatMed18:1003-1006.FrankelAD,PaboCO(1988)Cellularuptakeofthetatproteinfromhumanimmunodeficiencyvirus.Cell55:1189-1193.GreenM,LoewensteinPM(1988)Autonomousfunctionaldomainsofchemicallysynthesizedhumanimmunodeficiencyvirustattrans-activatorprotein.Cell55:1179-1188.HiroseH,TakeuchiT,OsakadaH,PujalsS,KatayamaS,NakaseI,KobayashiS,HaraguchiT,FutakiS(2012)Transientfocalmembranedeformationinducedbyarginine-richpeptidesleadstotheirdirectpenetrationintocells.MolTher20:984-993.HoA,SchwarzeSR,MermelsteinSJ,WaksmanG,DowdySF(2001)Syntheticproteintransductiondomains:enhancedtransductionpotentialinvitroandinvivo.CancerRes61:474-477.KacprzakMM,PeinadoJR,ThanME,AppelJ,HenrichS,LipkindG,HoughtenRA,BodeW,LindbergI(2004)Inhibitionoffurinbypolyarginine-containingpeptides:nanomolarinhibitionbynona-D-arginine.JBiolChem279:36788-36794.KilicKilicE,DietzGPHM(2003)IntravenousTAT-GDNFisprotectiveafterfocalcerebralischemiainmice.Stroke34:1304-1310.LaiY,DuL,DunsmoreKE,JenkinsLW,WongHR,ClarkRS(2005)Selectivelyincreasinginducibleheatshockprotein70viaTAT-proteintransductionprotectsneuronsfromnitrosativestressandexcitotoxicity.JNeurochem94:360-366.LiuX,M,PeiDS,GuanQH,SunYF,WangXT,ZhangQX,ZhangGY(2006)NeuroprotectionofTat-GluR6-9cagainstneuronaldeathinducedbykainateinrathippocampusvianuclearandnon-nuclearpathways.JBiolChem281:17432-17445.MeadeAJ,MeloniBP,MastagliaFL,KnuckeyNW(2009)Theapplicationofcellpenetratingpeptidesforthedeliveryofneuroprotectivepeptides/proteinsinexperimentalcerebralischemiastudies.JExpStrokeTranslMed2:22-40.MeadeAJMeloniBP,MastagliaFL,WattPMKnuckeyNW(2010a)AP-1inhibitorypeptidesattenuateinvitrocorticalneuronalcelldeathinducedbykainicacid.BrainRes1360:8-16.MeadeAJ,MeloniBP,CrossJBakkerAJ,FearMW,MastagliaFL,WattPM,KnuckeyNW(2010b)AP-1inhibitorypeptidesareneuroprotectivefollowingacuteglutamateexcitotoxicityinprimarycorticalneuronalculturesJNeurochem112:258-270.MeloniBP,MajdaBT,KnuckeyNW(2001)Establishmentofneuronalinvitromodelsofischemiain96-wellmicrotiterstrip-platesthatresultinacute,progressiveanddelayedneuronaldeath.Neuroscience108:17-26.MeloniBP,MeadeAJ,KitikomolsukD,KnuckeyNW(2011)Charaoterisationofneuronalcelldeathinacuteanddelayedinvitroischemia(oxygen/glucosedeprivation)models.JNeurosciMethods195:67-74.MillettiF(2012)Cell-penetratingpeptides:classes,origin,andcurrentlandscape.DrugDiscovToday17:850-860.MoschosSA,JonesSW,PerryMM,WilliamsAE,ErjefaltJS,TurnerJJ,BarnesPJ,SproatBS,GaitMJ,LindsayMA(2007)LungdeliverystudiesusingsiRNAconjugatedtoTAT(48-60)andpenetratinrevealpeptideinducedreductioningeneexpressionandinductionofinnateimmunity.BioconjugChem18:1450-1459.NagelF,FalkenburgerBH,L,KowskyS,C,SchulzJB,M,DietzGP(2008)Tat-Hsp70protectsdopaminergicneuronsinmidbrainculturesandinthesubstantianigrainmodelsofParkinson′sdisease.JNeurochem105:853-864.NathA,PsooyK,MartinC,KnudsenBMagnusonDS,HaugheyN,GeigerJD(1996)Identificationofahumanimmunodeficiencyvirustype1Tatepitopethatisneuroexcitatoryandneurotoxic.JVirol70:1475-1480.Palm-ApergiC,P,DowdySF(2012)Docell-penetratingpeptidesactually″penetrate″cellularmembranes?MolTher20:695-697.VaslinA,RummelC,ClarkePG(2009)UnconjugatedTATcarrierpeptideprotectsagainstexcitotoxicity.NeurotoxRes15:123-126.XuW,ZhouM,BaudryM(2008)NeuroprotectionbycellpermeableTAT-mGluR1peptideinischemia:synergybatweencarrierandcargosequences.Neuroscientist14:409-414.ZhangS,TaghibiglouC,GirlingK,DongZ,LinSZ,LeeW,ShyuWC,WangYT(2013)CriticalroleofincreasedPTENnucleartranslocationinexcitotoxicandischemicneuronalinjuriesJNeurosci33:7997-8008.當前第1頁1 2 3